
Многоплановая композиция: Взаимодействие планов. Многослойные кадры / Fotoshkola.net
Взаимодействие планов. Многослойные кадры / Fotoshkola.net
«Есть фотографии, которые могут не только сохранить удивительный выразительный эффект присутствия, благодаря редкому элементу или необычной композиции, но способны излучать особенную атмосферу сами по себе».
Робер Дуано
Глубинная композиция в репортаже
Многослойные, глубинные композиции занимают особое место среди репортажных снимков.
Я уже обращал ваше внимание на то, что ситуацию или человека можно снять в одной плоскости, лаконично, плакатно, исключив из кадра лишние детали. Такие фотографии легко читаются и хорошо смотрятся на обложке журналов, газетной полосе, главной странице интернет-сайта, с них можно начинать любой видеоряд.
Вместе с тем, событие, которое вы снимаете, разворачивается в пространстве. Что-то может происходить перед главным объектом съемки или за ним в отдалении, то есть на переднем или на заднем планах. Эти объекты, связанные общим смыслом, можно включить в кадр и заставить их «говорить» вместе. Причем, объекты или ситуации второго плана способны глубже раскрыть смысл происходящего, нести в себе подтекст, подталкивать к аналогии, контрастировать с главным объектом и тем самым подчеркивать его собственные свойства.
Эти объекты, связанные общим смыслом, можно включить в кадр и заставить их «говорить» вместе. Причем, объекты или ситуации второго плана способны глубже раскрыть смысл происходящего, нести в себе подтекст, подталкивать к аналогии, контрастировать с главным объектом и тем самым подчеркивать его собственные свойства.
Кадр, в котором показывается событие в разных плоскостях пространства, называется многоплановым или многослойным, а композицию такого снимка можно назвать глубинной.
Глубинные композиции кинематографичны, то есть напоминают сцену из фильма. В них всегда есть передний план, не обязательно являющийся главным. Многослойные кадры объемны, в них хочется всматриваться. Общее впечатление, идея снимка создаются всеми деталями, информационным и художественным взаимодействием планов. Удачный многоплановый снимок — это целый рассказ в одном кадре, в нем все важно до мелочей.
Рассмотрим примеры:
Анри Картье-Брессон. Рабочий баржи на Сене (Франция, 1957).
Павел Кривцов. Весна на о. Кижи
Весна на о. Кижи
В представленных фотографиях Павла Кривцова и Анри Картье-Брессона трудно выделить главный объект, настолько все слои этих снимков связаны между собой в художественном и повествовательном смыслах. Но если мы будем анализировать композицию, то увидим типичную глубинную структуру с распределением сюжетно значимых объектов на переднем и заднем планах. У Картье-Брессона это ситуация взаимоотношения рабочего (передний план) и его семьи (задний план). У Кривцова — на переднем плане телега, картошка и дети, на заднем плане известная древняя деревянная церковь. Они создают единый образ, который и обозначен в названии фотографии — «Весна на Кижах».
Репортажные снимки, созданные по принципу многоплановой композиции, очень эффектны, но получить хороший снимок-рассказ непросто. В арсенале даже опытного репортера найдется немного таких фотографий, и их наличие — свидетельство мастерства. Я имею в виду сложные многоплановые кадры, в которых запечатлено взаимодействие сцен, а не простое расположение объектов в разных плоскостях.
Как создаются многослойные кадры
Рождаются глубинные композиции в результате активного поиска, внимательного наблюдения, удачного стечения обстоятельств и изобретательности фотографа. На практике важны оба навыка — умение видеть и строить композицию.
Правильно выстроенный снимок с использованием элементов постановки будет выглядеть естественным, «случайным». «За кадром» останутся усилия фотографа — поиск точки съемки, управление героем, выжидание момента, когда планы сольются в единый образ. Для фотографа важно как бы предчувствовать этот момент и замечать все, что происходит вокруг героя съемки. К такого рода выстроенным кадрам можно отнести фотографию Павла Кривцова «Поэт-пастух из Белгородской области Владимир Михалев».
Павел Кривцов во время съемки, безусловно, имел возможность приглядеться, обдумать и построить этот кадр. В фотографии несколько слоев: от котелка на переднем плане до закатного неба на дальнем. Все элементы пространства связаны общим настроением, созвучным душевному состоянию героя. Эффект нашего присутствия создает «редкий элемент» (Роббер Дуано) и организация объектов снимка.
Эффект нашего присутствия создает «редкий элемент» (Роббер Дуано) и организация объектов снимка.
Аналогичным образом был построен мой снимок «Свидание с матерью». В колонии, в камере для свиданий, мне нужно было снять бывшего солдата, осужденного за несуществующее военное преступление в Афганистане. Милиционера я деликатно «разместил» в дверях камеры. Во время съемки одновременно следил за двумя объектами: матерью с сыном и милиционером. Взаимоотношения матери с сыном, их взгляды, жесты и поза милиционера в одно прекрасное мгновение «созрели», и кадр был сделан! Фигура милиционера на заднем плане рассказывает нам о месте, где происходит встреча и усиливает драматизм ситуации.
Яркой иллюстрацией, где оба плана работают на одну мысль — сострадание к ребенку перед операцией, является репортажная фотография «Все будет хорошо». Это не постановка. Соединить два плана я успел, едва-едва отскочив в угол операционной комнаты для того, чтобы включить в кадр хирурга, натягивающего резиновые перчатки. Я вжался в угол, и фиксированный объектив (кажется, с фокусным расстоянием 135 мм) не позволил мне включить в кадр большую часть фигуры хирурга.
Я вжался в угол, и фиксированный объектив (кажется, с фокусным расстоянием 135 мм) не позволил мне включить в кадр большую часть фигуры хирурга.
В лучших сложносочиненных репортажных кадрах кристаллизируются все способности фотографа как художника, мыслителя, конструктора и исследователя. Но все же путь к «решающему моменту», и я не устану об этом повторять, начинается с умения видеть красоту, трагизм или необычность, контраст или согласованность жизненных ситуаций. Посмотрим, как это умение видеть передается в других многоплановых кадрах.
Павел Кривцов. Ветераны войны села Койнас Архангельской области (из серии).
Павел Кривцов. Лена Кочеткова — художник декоративной росписи из Жостово.
Фото Геннадия Михеева
Фото Геннадия Михеева.
Фото Александры Рогозиной.
Простые решения в многослойной композиции
В обычной репортажной съемке довольно легко даются простые многоплановые композиции, если вы развиваете в себе глубинное зрение. То есть не забываете наблюдать за объектами и ситуациями второго плана, происходящими рядом с главным героем вашей съемки.
Рассмотрим первый вариант простого решения в многослойной композиции.
Выбором точки съемки или искусственно постановкой, расположите на переднем плане в кадре какой-либо предмет (или предметы), с которым связана вся фотографируемая ситуация. За этим предметом (вещью) будет разыгрываться основная сцена съемки. Сделайте снимок, фокусируясь на главной ситуации.
Примеры:
Фото Александры Рогозиной.
Михаил Рогозин. Парижский дом моды Lanvin (из серии).
Михаил Рогозин. В Соловецкой обители (из серии).
Фото Геннадия Михеева.
Александра Рогозина. Мастер-класс по анимации (из серии).
Александра Рогозина. Мастер-класс по анимации (из серии).
Чем интересны простые глубинные «предметные» композиции? Во-первых, они создают пространственное чувство, создают эффект присутствия. Во-вторых, вынесением предмета на передний план вы привлекаете к нему внимание, позволяете почувствовать его фактуру и рассмотреть его вблизи. В-третьих, из поля зрения не уходит само действие, ситуация или человек, с которым как-то связан предмет переднего плана.
Второй вариант простой многослойной композиции — это взаимодействие сцен, ситуаций.
Под сценой, ситуацией я понимаю взаимодействие, занятия людей. Но даже один человек, поглощенный своим делом, уже составляет сцену, подобно мизансцене, игре одного актера. Распределение сцен переднего и заднего планов в кадре составляет суть сложносочиненных сценических кадров, которые были рассмотрены выше.
В простом варианте добротный, беспроигрышный «сценический» многоплановый кадр можно создать, искусственно включая на передний план фигуру или фрагменты фигуры второстепенного героя, например, часть головы, плеча, руки. Фокусировкой (наводкой резкости) выделите главного героя. Такой кадр будет отличаться максимальной емкостью, информационной насыщенностью.
Примеры:
Фото Геннадия Михеева.
Фото Александры Рогозиной.
Фото Александры Рогозиной.
Фото Михаила Рогозина.
Александра Рогозина. На мастер-классе по анимации (вся серия в примерах).
В заключение урока дам совет: не эксплуатируйте прием построения простой многоплановой композиции без нужды, по всякому поводу. Помните, что это всего лишь один из фотографических приемов. Перебор таких фотографий в фотосерии усложняет ее восприятие, может сделать ее однообразной. Но один-два кадра помогут вам раскрыть тему фотоистории глубже.
Помните, что это всего лишь один из фотографических приемов. Перебор таких фотографий в фотосерии усложняет ее восприятие, может сделать ее однообразной. Но один-два кадра помогут вам раскрыть тему фотоистории глубже.
Дополнительная литература по теме урока
- Дыко Лидия. «Беседы о фотомастерстве», М., «Искусство», 1977
- Медынский Сергей. «Компонуем кинокадр», М., «Искусство», 1992
- Официальные партнёры
7 приемов композиции, которые фотографы могут заимствовать из фильмов
октября 1, 2020
Кино — бесценный источник вдохновения и идей для фотографа. Там можно найти огромное количество приемов для своих снимков: свет, композиция, колористика, стиль, работа с актером как с моделью.
Настя Одинцова, семейный фотограф и соавтор курса «Дети в фокусе», поделилась с нами своими любимыми фильмами и рассказала о «киношных» приемах композиции, которые помогут сделать фотографии яркими и интересными. Уверены, что после этой статьи вы будете смотреть фильмы совершенно под другим углом.
Уверены, что после этой статьи вы будете смотреть фильмы совершенно под другим углом.
Я часто смотрю кино с точки зрения фотографа и анализирую композицию, свет, цвета. Это помогает развивать насмотренность и подбирать гармоничный гардероб, подмечать места для съемок, использовать цвет и линии в кадре.
В этой статье я собрала кадры из пяти моих любимых фильмов: «Амели», «Свет в океане», «Невероятная жизнь Уолтера Митти», «Девушка из Дании» и «Поезд на Дарджилинг. Отчаянные путешественники». Их создали абсолютно разные режиссеры, но их объединяет одно — классная композиция. Давайте вместе попробуем посмотреть на нее глазами фотографа.
Многоплановость
В фотографии принято выделять передний, средний и задний план. Когда снимок многоплановый, это создает объем и зритель может глубже погрузиться в кадр. Обычно это достигается благодаря широкоугольной оптике и закрытой диафрагме. Многие фотографы не любят использовать такие объективы, потому что на фотографиях появляется много «мусора» — гораздо проще спрятать все ненужное за красивым боке. Однако часто именно за счет многоплановой композиции в кадре рождается сюжет, показывается объем и создается история.
Однако часто именно за счет многоплановой композиции в кадре рождается сюжет, показывается объем и создается история.
Посмотрите, как на этих стоп-кадрах освещены персонажи. Несмотря на то, что герои освещены даже на дальних планах, акценты расставлены так, что главный герой — это композиционный центр кадра — отлично считывается.
Многоплановые кадры из фильмов «Невероятная жизнь Уолтера Митти» и «Девушка из Дании»
Самое главное правило многоплановости — планы не должны перекрывать друг друга. Посмотрите, как расположены герои на этих стоп-кадрах. Как они стоят? Почему именно так? Оказывается, здесь нет случайностей: все персонажи стоят идеально и ни один из них не перекрывает другого. Получить такой кадр можно при закрытой диафрагме, чтобы прорисовать все планы.
Многоплановые кадры из фильмов «Поезд на Дарджилинг. Отчаянные путешественники» и «Амели»
Голландский угол
Голландский угол — это намеренный завал линии горизонта. При этом важно, чтобы на фото были вертикальные линии. Они придают композиции устойчивость, поэтому кадр перестает визуально «падать». Если вы случайно сняли немного криво, не спешите удалять снимок и сначала проанализируйте: возможно, заваленный горизонт сделал фотографию более эффектной.
Они придают композиции устойчивость, поэтому кадр перестает визуально «падать». Если вы случайно сняли немного криво, не спешите удалять снимок и сначала проанализируйте: возможно, заваленный горизонт сделал фотографию более эффектной.
Голландский угол помогает создать динамику в кадре. Его часто используют, чтобы разнообразить диалоговые сцены или передать какое-то напряжение у героя. Например, так этот прием использовали в фильме «Амели».
Голландский угол в кадрах из фильма «Амели»
Фрейминг
Хороший способ выделить композиционный центр и акцентировать внимание на объекте съемки — использовать фрейминг. В переводе с английского «фрейминг» значит «обрамление». Другими словами, это рамка.
Естественные и искусственные рамки можно найти везде. Например, на стоп-кадрах ниже это окна поезда и дверной проем.
Кадры с рамками из фильмов «Поезд на Дарджилинг. Отчаянные путешественники» и «Свет в океане»
Рамка может быть как на переднем, так и на заднем плане. При этом она даже не обязательно должна быть цельной. Это могут быть и несколько предметов, которые зрительно образуют рамку.
При этом она даже не обязательно должна быть цельной. Это могут быть и несколько предметов, которые зрительно образуют рамку.
Посмотрите на кадры из фильмов. В них используются разные варианты рамок на переднем плане. На первом стоп-кадре это оконный проем. Здесь сразу две рамки, которые визуально делят кадр пополам. Это намекает зрителю, что люди не вместе и их дороги скорее всего уже не сойдутся. А на втором кадре модель снята внутри зеркала. Благодаря фреймингу композиционный центр воспринимается как картина, висящая на стене.
Кадры с рамками из фильмов «Девушка из Дании» и «Амели»
Симметрия
Симметрия — это мощный прием композиции, который подсмотрен у природы. Вы каждый день видите вокруг себя симметрию: вертикальная — тело человека, горизонтальная — отражение в озере, радиальная — цветок или снежинка.
Если вы хотите прокачать свои фотографии потрясающе ровной симметрией, то вам обязательно нужно посмотреть фильмы Уэса Андерсона. В них локации настолько утрированы своей симметричностью, что это выглядят просто потрясающе. Ниже несколько кадров из фильма Уэса Андерсона «Поезд на Дарджилинг», но я советую обязательно посмотреть еще и киноленту «Отель „Гранд Будапешт“» — там «симметрия еще более симметрична».
Ниже несколько кадров из фильма Уэса Андерсона «Поезд на Дарджилинг», но я советую обязательно посмотреть еще и киноленту «Отель „Гранд Будапешт“» — там «симметрия еще более симметрична».
Симметрия в фильме «Поезд на Дарджилинг. Отчаянные путешественники»
Не всегда симметрия должна быть точной, как отражение в зеркале. Равновесие может быть задано схожими элементами в противоположных частях фотографии. Посмотрите на примеры ниже. Здесь два стоп-кадра, части которых имеют явные различия. Однако, они выглядят очень симметрично.
Особенно интересен второй скриншот. Часто в симметрии композиционный центр находится по центру кадра. Благодаря отражающему предмету, который находится перед камерой, у главного героя появляются интересные отражения. Похожего эффекта можно добиться, если приставить к объективу предмет с отражающей поверхностью. Например, для плотного и контрастного отражения подойдет экран мобильного телефона, а для полупрозрачного — стекло.
Симметрия в фильмах «Девушка из Дании» и «Свет в океане»
Направление взгляда
Часто в фильмах оставляют воздух и пространство для взгляда героев. Так в кадре появляется сильная эмоция. А иногда это правило наоборот нарушается. Например, режиссер Том Хупер в фильме «Девушка из Дании» экспериментирует с широкоугольной оптикой и располагает героев критично близко к границам кадра. Посмотрите на второй стоп-кадр: взгляд героя будто упирается в незримую границу.
Так в кадре появляется сильная эмоция. А иногда это правило наоборот нарушается. Например, режиссер Том Хупер в фильме «Девушка из Дании» экспериментирует с широкоугольной оптикой и располагает героев критично близко к границам кадра. Посмотрите на второй стоп-кадр: взгляд героя будто упирается в незримую границу.
Этот прием позволяет играть с пространством. Представьте человека, который куда-то идет. Если мы оставим больше места по направлению движения, появится ощущение, что человек приближается к месту назначения. А если оставим больше пространства позади героя, будет казаться, что он отдаляется от своей цели.
Разное направление взгляда в кадрах из фильмов «Невероятная жизнь Уолтера Митти» и «Девушка из Дании»
Выбор цветов
Цвет — один из самых очевидных элементов композиции. Его постоянно используют в кино, чтобы привлечь внимание, расставить акценты или создать нужное настроение. Я выделила несколько способов работы с цветом, которые можно использовать.
Цветовая палитра
Часто в фильмах выбирают определенную цветовую гамму, которая держится на протяжении всей киноленты. Например, в фильме «Невероятная жизнь Уолтера Митти» три основных цвета: красный, синий и желтый. Они выдержаны везде, даже в натюрморте с обычной едой в самолете.
Например, в фильме «Невероятная жизнь Уолтера Митти» три основных цвета: красный, синий и желтый. Они выдержаны везде, даже в натюрморте с обычной едой в самолете.
Три основных цвета в фильме «Невероятная жизнь Уолтера Митти»
Иногда цвет помогает передать мысль и замысел намного глубже. Посмотрите на стоп-кадр ниже. Здесь отсылка к фильму «Матрица», где главный герой выбирал пилюлю красного цвета, потому что синий означал конец пути.
Использование цвета в фильме «Невероятная жизнь Уолтера Митти»
Монохром
Многие ошибочно считают монохромными черно-белые фотографии. На самом деле в монохроме за основу берется один цвет и его оттенки — тогда фотография обычно получается мягкой и гармоничной. Чтобы получить монохромную картинку, можно специально одевать моделей в одежду нужного цвета, тщательно настраивать баланс белого на камере и т.д. Такой композиционный прием помогает зрителю сосредоточиться на происходящем, не отвлекаясь на детали.
Монохромные кадры из фильмов «Поезд на Дарджилинг. Отчаянные путешественники» и «Амели»
Отчаянные путешественники» и «Амели»
Цветовой акцент
Любимый прием многих фотографов и операторов — цветовой акцент. Это яркое цветовое пятно, которое приковывает взгляд. Обычно так выделяют главный объект съемки.
Посмотрите на кадры из фильма «Амели». Вся картина снята в трех основных цветах: желтый, красный и зеленый. Однако на некоторых кадрах присутствуют вкрапления ярко-синего. Это холодный цвет, который явно доминирует на фоне теплых оттенков. Такой акцент может показаться спорным, ведь синие предметы не главные в кадре. Но я бы сказала, что это небольшое нарушение правил — выход за границы привычного, которое имеет место быть в любом творчестве.
Цветовые акценты в фильме «Амели»
Направляющие линии
Направляющие линии — это еще один способ сделать акцент. Они ведут в композиционный центр и играют огромную роль в том, как мы воспринимаем образ. При этом они могут быть разными. Вертикальные обычно создают динамику в кадре, а горизонтальные настолько привычны, что дарят ощущение покоя и стабильности.
Для примера я отобрала необычные варианты использования направляющих линий. В кадре из фильма «Амели» сразу несколько линий, направляющих взгляд к человеку. И самая главная из них — стрелка со словом «toilettes». Это бесподобный прием, который не только указывает нам на главного героя, но и раскрывает информацию о нем. И действительно, в фильме этот персонаж несет отрицательную роль: у него грязный рот, и направляющая линия нам на это указывает.
Во втором кадре нет явных направляющих, но они создаются благодаря линиям взгляда людей, расположенных по бокам от композиционного центра. Одновременно герой выделен светом, что усиливает акцент.
Использование направляющих линий в фильмах «Амели» и «Невероятная жизнь Уолтера Митти»
Конечно, в кино еще много композиционных приемов, которые можно перенести в фотографию. Каждый режиссер и оператор видит композицию по-своему, поэтому не стоит ограничиваться этими пятью фильмами. Я рекомендую еще несколько кинокартин, которые помогут развивать насмотренность:
- «Королевство полной луны»,
- «Отель „Гранд Будапешт“»,
- «Соль Земли»,
- все фильмы Андрея Тарковского.


Мне хочется, чтобы после просмотра вы научились смотреть фильмы под другим углом, замечать свет, цвет, композицию. И, конечно, использовать эти приемы в своих фотографиях.
Еще кое-что интересное
Полезные и вдохновляющие статьи 2 раза в месяц.
Без спама и только по делу
Композиционные приемы
- Главная
- Композиционные приемы
Композиция — это расположение элементов в художественном произведении.
Хотя в общем смысле любой фрагмент музыки, живописи или скульптуры можно назвать композицией, термин обычно относится к расположению элементов в произведении искусства. Это то, как элементы искусства и дизайна, линия, форма, цвет, значение, текстура, форма и пространство — организованы или составлены в соответствии с принципами искусства и дизайна. В классической традиции использовались треугольные или пирамидальные композиции, потому что они создавали ощущение равновесия и гармонии, устраивая фигуры в устойчивую общую геометрическую структуру. Это можно увидеть, например, в грубо конической группировке животных в «Mares and Foals» Джорджа Стаббса
.Джордж Стаббс. «Mares and Foals»
Идея композиции как корректировки отношений элементов внутри границы холста оставалась неоспоримой благодаря появлению ранних современных движений, таких как кубизм и абстрактное искусство.
Композиция по словам Анри Матисса
«Композиция — это искусство декоративного оформления разнообразных элементов картины, чтобы выразить свои чувства».
 — Анри Матисс в «Заметках художника».
— Анри Матисс в «Заметках художника». Элементы композиции
Элементы, принципы искусства и дизайна являются основой языка, на котором мы говорим об искусстве. Элементами искусства являются визуальные инструменты, которые художник использует для создания композиции. Это линия, форма, цвет, значение, форма, текстура и пространство.
Хотите научиться рисовать? Онлайн школы рисования.
Разновидности композиционных решений показаны на рисунках ниже.
Формат
Пространство
Одноплановая композиция / Многоплановая композиция
Композиционный центр
В центре / Смещен от центра
Симметрия
Симметрия / Асимметрия
Равновесие
Устойчивое / Неустойчивое
Динамика
Движение / Статика
Ритм / Контраст
Светотень (объемность)
Композиция
Открытая / Закрытая
Компоновка
Равновесие
Пропорциональное / Стесненное
Расположение
В центре — уравновешенность / В углу — спокойствие (желание переместить)
Ниже оси — тяжесть / Выше оси — легкость
Ритмы
Тональный ритм / Цветовой ритм
Тональный и цветовой ритмы в изображении
Понравилась статья? Поделитесь с друзьями.

Твитнуть
Поделиться
Поделиться
Отправить
Класснуть
Линкануть
- Назад
- Вперед
ПОЛУЧИ ПРОФЕССИЮ: ХУДОЖНИК ПЕСОЧНОЙ АНИМАЦИИ
Получите востребованную профессию дизайнера
Хотите научиться рисовать? Онлайн школы рисования.
Идеальный курс для новичков!
Учимся работать сразу с 5-ю художественными материалами,
Посмотреть
- Графические программы для дизайнеров и архитекторов
- Композиция картины
- Рисунок гипсовой головы
- Техники и технологии акварельной живописи
- Технологии живописи
- Русский художник Рерих
- Чистка и лакировка картин
- Картины в интерьере
- Уход за картинами
 org/Article»>
Андрей Рублев
org/Article»>
Андрей РублевПрисоединяйтесь !
Сайт с открытками
Где и как научиться рисовать
Компьютерная графика онлайн
Руководство по программированию цепочки буферов компоновки — Win32 apps
- Статья
- Чтение занимает 13 мин
API подкачки композиции является духовным преемником цепочки буферов DXGI, что позволяет приложениям отображать и представлять содержимое на экране. Существует несколько преимуществ использования этого API по сравнению с цепочкой буферов DXGI. Более точное управление предоставляется вашему приложению относительно состояния цепочки буферов, и больше свободы предоставляется, когда дело доходит до того, как используется цепочка буферов. Кроме того, API предоставляет лучшую историю для точного текущего времени.
Существует несколько преимуществ использования этого API по сравнению с цепочкой буферов DXGI. Более точное управление предоставляется вашему приложению относительно состояния цепочки буферов, и больше свободы предоставляется, когда дело доходит до того, как используется цепочка буферов. Кроме того, API предоставляет лучшую историю для точного текущего времени.
Что такое презентация?
Презентация — это концепция отображения результатов операций рисования на экране. Настоящее представление представляет собой один экземпляр презентации — запрос на отображение результатов операции рисования на одном буфере на экране. В настоящее время могут содержаться дополнительные атрибуты, описывающие отображение на экране. В этом API может также иметь целевое время, которое является системной меткой времени (время прерывания), описывающее идеальное время, которое должно отображаться в настоящее время. Приложение может использовать его для более точного контроля скорости отображения содержимого на экране и синхронизации представлений с другими событиями в системе, такими как звуковая дорожка.
В основе презентации лежит синхронизация. То есть операции рисования обычно выполняются GPU, а не ЦП, и, как таковое, они выполняются на асинхронной временной шкале из ЦП, выдавшей операции изначально. Презентация — это операция, отправленная на GPU, которая гарантирует, что операции рисования, выпущенные ранее, будут завершены до отображения буфера на экране.
Приложение обычно выдает много подарков с течением времени и имеет несколько текстур, которые можно выбрать при выдаче подарков. Приложение должно использовать механизмы синхронизации, предоставляемые этим API, чтобы убедиться, что после рисования и представления буфера вы не будете рисовать в этот буфер еще раз, пока это не будет показано и впоследствии заменено новым буфером из последующего текущего. В противном случае содержимое буфера, которое должно быть представлено приложением, может быть перезаписано по мере отображения на экране.
Режимы презентации — композиция, многоплановая наложение и iflip
Буферы, представленные приложением, могут отображаться системой несколькими разными способами.
Самый простой способ, который является значением по умолчанию, заключается в том, что настоящее представление будет отправлено в DWM, а DWM будет отображать кадр на основе представленного буфера. То есть есть есть копия (или более точно, трехмерная отрисовка) буфера презентации в обратный буфер, который DWM отправляет на экран. Этот метод отображения текущего объекта называется Composition.
Более производительным режимом отображения текущего представления будет сканирование буфера презентации непосредственно на оборудование и исключение выполняемой копии. Этот метод отображения текущего объекта называется прямым сканированием. При обработке презентаций DWM может решить запрограммировать оборудование для прямого сканирования буфера презентации, назначив буфер многоплановой плоскости наложения (или плоскость MPO, короче говоря), или перевернуть буфер на оборудование (называемое прямым переворачиванием).
Еще более производительный способ отображения презентации будет отображаться непосредственно ядром графики и полностью обходить DWM. Этот метод представления называется независимым отражением (iflip). И многоплановый наложение, и если см. в разделе «Для оптимальной производительности», используйте модель пролистывания DXGI.
Этот метод представления называется независимым отражением (iflip). И многоплановый наложение, и если см. в разделе «Для оптимальной производительности», используйте модель пролистывания DXGI.
Композиция является наиболее легко поддерживаемой, но и наименее эффективной. Поверхность должна быть специально выделена, чтобы иметь право на прямое сканирование или iflip, и этот тип специального выделения имеет более строгие требования к системе, чем цепочка буферов композиции. Она доступна только на оборудовании WDDM 3.0 и более поздней версии. В результате приложение может запрашивать поддержку API для презентации только для композиции, а также презентацию, которая квалифизируется для прямого сканирования или iflip.
Фабрика презентаций, возможность проверки и руководитель презентаций
Первым объектом, который приложение будет использовать из API подкачки композиции, является фабрика презентаций. Фабрика презентаций создается приложением и привязывается к устройству Direct3D, которое приложение передает в вызов для создания, и, таким образом, имеет сходство с видеоадаптером, связанным с этим устройством.
Фабрика презентаций предоставляет методы для проверки того, может ли текущая система и графическое устройство использовать API буферной цепочки композиции. Для проверки поддержки системы можно использовать такие методы возможностей, как IPresentationFactory::IsPresentationSupported . Если методы возможностей указывают системную поддержку API, можно использовать фабрику презентаций для создания диспетчера презентаций. Этот диспетчер презентаций — это объект, используемый для выполнения функций презентации, и привязан к тому же устройству Direct3D и видеоадаптеру, что и фабрика презентаций, которая использовалась для его создания.
В настоящее время требования к системе для использования API цепочки буферов композиции — это драйверы GPU, поддерживающие WDDM (модель драйвера устройств Windows) 2.0, и Windows 11 (сборка 10.0.22000.194) или более поздней версии. Чтобы использовать API цепочки буферов композиции наиболее производительным способом (прямой проверочный и независимый пролистывание, или если скольжение), системам потребуются драйверы GPU, поддерживающие WDDM 3. 0.
0.
Если система не может использовать API цепочки буферов композиции, приложению потребуется отдельный путь к коду для обработки презентации с помощью старых методов, таких как цепочка буферов DXGI.
Регистрация буферов презентации для представления
Диспетчер презентаций отслеживает буферы, которые он может представить. Чтобы представить текстуру Direct3D, приложение должно сначала создать ее с помощью Direct3D, а затем зарегистрировать ее в диспетчере презентаций. Если текстура зарегистрирована в диспетчере презентаций, она называется буфером презентации и может с этого момента отображаться на экране этим диспетчером презентаций. Приложение может добавлять и удалять буферы презентаций по своему усмотрению, хотя существует максимальное количество буферов презентации, которые можно добавить в один диспетчер презентаций (в настоящее время 31). Эти буферы презентации также могут иметь различные размеры и форматы, которые вступают в силу в качестве отдельного буфера презентации.
Текстуру можно зарегистрировать с любым количеством руководителей презентаций; однако в большинстве случаев это не считается нормальным использованием и будет приводить к сложным сценариям синхронизации, которые ваше приложение будет отвечать за управление.
Определение содержимого для представления
Как правило, буферы, которые мы представляем, должны быть связаны с содержимым в визуальном дереве. Поэтому нам нужно определить вид привязки , чтобы при появлении проблем с приложением было ясно, где в визуальном дереве представлены буферы. Мы называем эту привязку содержимым презентации.
Представленное содержимое может принимать множество форм. Ваше приложение может захотеть представить один буфер для отображения или представить стереоконтентное содержимое с буферами для левого и правого глаз и т. д. Начальная версия этого API обеспечивает поддержку представления одного буфера на экране.
Мы определяем поверхность презентации как форму содержимого презентации, в которую одновременно предоставляется один буфер. Область презентации может быть задана в виде содержимого в визуальном дереве и может одновременно отображать один буфер презентации на экране. Диспетчер презентаций обновит буфер, отображаемый одной или несколькими поверхностями презентации атомарным образом.
Область презентации может быть задана в виде содержимого в визуальном дереве и может одновременно отображать один буфер презентации на экране. Диспетчер презентаций обновит буфер, отображаемый одной или несколькими поверхностями презентации атомарным образом.
Диспетчер презентаций можно использовать для создания одной или нескольких поверхностей презентации для заданного дескриптора поверхности композиции. Каждый дескриптор поверхности композиции можно привязать к одному или нескольким визуальным элементам в визуальном дереве (стратегиями, описанными в Windows. ПОЛЬЗОВАТЕЛЬСКОГО ИНТЕРФЕЙСА. Документация по API composition и DirectComposition) для определения связи между связанной поверхностью презентации и местом ее отображения в визуальном дереве. Приложение может обновить одну или несколько поверхностей презентации, которые отправляются в систему и выполняются при следующей текущей операции.
Обратите внимание, что диспетчер презентаций может представить любой буфер презентации любому количеству поверхностей презентации, которые он хочет. Ограничения отсутствуют. Однако приложение может отслеживать, какие буферы вы выпустили, и где, чтобы убедиться, что вы не пытаетесь выдать новый рисунок в этот буфер, пока он по-прежнему отображается поверхностью презентации.
Ограничения отсутствуют. Однако приложение может отслеживать, какие буферы вы выпустили, и где, чтобы убедиться, что вы не пытаетесь выдать новый рисунок в этот буфер, пока он по-прежнему отображается поверхностью презентации.
Применение свойств к поверхности презентации
Помимо указания буферов, отображаемых в поверхности презентации, может также указывать различные другие свойства для этой области презентации. К ним относятся свойства, определяющие способ выборки исходной текстуры, включая альфа-режим и цветовое пространство, способ преобразования и размещения исходной текстуры, а также любые отображаемые или ограничения обратного чтения для защищенного содержимого. Все они предоставляются в качестве методов задания свойств на поверхности презентации, которые могут быть изменены приложением, и, как и обновления буфера, вступают в силу при наличии приложения.
Презентация в презентации
После создания поверхностей презентации приложение регистрирует буферы презентаций и указывает обновления, которые будут выдаваться во время текущего представления, можно применить эти свойства. Ваше приложение выдает презентацию через диспетчер презентаций. При обработке системой все обновления применяются атомарно. Кроме того, приложение может также указать другие свойства текущего объекта, такие как идеальное время ( текущее целевое время ) и другие редкие свойства, такие как предполагаемая частота содержимого, которые можно использовать для включения пользовательских режимов обновления в системе. Так как представление может быть запланировано в определенное время, приложение может выдавать несколько представлений заранее. Эти подарки будут обрабатываться по одному по мере достижения запланированного времени.
Ваше приложение выдает презентацию через диспетчер презентаций. При обработке системой все обновления применяются атомарно. Кроме того, приложение может также указать другие свойства текущего объекта, такие как идеальное время ( текущее целевое время ) и другие редкие свойства, такие как предполагаемая частота содержимого, которые можно использовать для включения пользовательских режимов обновления в системе. Так как представление может быть запланировано в определенное время, приложение может выдавать несколько представлений заранее. Эти подарки будут обрабатываться по одному по мере достижения запланированного времени.
Синхронизация презентации
Приложение должно быть уверенным, что при отрисовке в буферы и проблемах выбирается буфер для отрисовки, на который в настоящее время не ссылается любой другой выдающийся предыдущий текущий момент, так как это может перезаписать содержимое буфера, которое они представляют. Кроме того, если приложение выдает отрисовку в буфер, который в настоящее время отображается поверхностью презентации в аппаратном обеспечении сканирования, то ее отрисовка может быть остановлена на неопределенный срок, так как Direct3D запрещает отрисовку переднего буфера.
API цепочки буферов композиции предоставляет несколько различных механизмов, позволяющих приложению практиковать правильную синхронизацию представленных буферов.
Считается, что буфер доступен , если отсутствуют выдающиеся представления, которые ссылаются на него, и в настоящее время он не отображается системой. В противном случае он недоступен. API предоставляет событие для каждого буфера презентации, указывающее, доступен ли буфер. Это самый простой метод синхронизации для используемого приложения. Перед рисованием в буфер и его отображением приложение может убедиться, что его доступное событие будет сигнализировать. Доступное событие для определенного буфера становится незаписанным, когда оно было привязано к поверхности презентации в API, и остается без знака вплоть до тех пор, пока настоящее время не станет устаревшим.
Во-вторых, руководитель презентации отслеживает единый выступающий забор для связи с приложением, которое было завершено. Значение ограждения соответствует настоящему идентификатору последнего текущего, который начал фазу прекращения жизненного цикла, как описано в разделе жизненного цикла ниже. После того как подарок переходит на этот этап, можно предположить, что все буферы, которые были заменены последующими представлениями, можно повторно использовать.
Значение ограждения соответствует настоящему идентификатору последнего текущего, который начал фазу прекращения жизненного цикла, как описано в разделе жизненного цикла ниже. После того как подарок переходит на этот этап, можно предположить, что все буферы, которые были заменены последующими представлениями, можно повторно использовать.
Этот метод синхронизации является более сложным, но обеспечивает больший контроль над регулированием рабочего процесса и является более информативным в отношении состояния системы в отношении глубины текущей текущей очереди. Общие сведения о жизненном цикле настоящего продукта см. в разделе ниже.
Жизненный цикл презентации
Представленные презентации диспетчера презентаций помещаются в очередь в систему как часть ее текущей очереди. Системные процессы представлены в порядке очередей. Кроме того, каждый подарок имеет уникальный (для руководителя презентации) связанный идентификатор, который является добавочным значением, назначенным настоящему, начиная с 1 для первого представления, и увеличивается на 1 для каждой последующей презентации. Этот идентификатор используется в различных частях API, таких как примитивы синхронизации и статистика представления, для ссылки на этот конкретный момент.
Этот идентификатор используется в различных частях API, таких как примитивы синхронизации и статистика представления, для ссылки на этот конкретный момент.
Каждый из них указывает, что проблемы с приложением соответствуют определенному жизненному циклу, как описано здесь.
После того как приложение настроит изменения, внесенные в качестве части презентации, оно будет использовать диспетчер презентаций для фактической выдачи настоящего. На этом этапе, как говорят, настоящее ожидается.
После ожидания презентация будет находиться в текущей очереди руководителя презентации, где она останется до тех пор, пока не произойдет одно из двух вещей.
- Настоящее становится отменено. Диспетчер презентаций позволяет приложению отменить ранее выданные подарки. Если это произойдет, то настоящее, как говорят, будет отменено, а затем он сразу же становится отставным. На этом переходе будут обновлены связанные события буфера, доступные для отмененного представления, однако текущее устаревшее ограждение не будет сигнализировать, так как отображаемый ранее (до отмененных представлений) останется отображаемым.
- Если это не отменено, настоящее время в конечном итоге становится готовым к обработке. Чтобы быть готовым, необходимо выполнить два основных условия.
- Все трудозатраты рисования, выданные контексту Direct3D до вызова настоящего, должны быть завершены. Это гарантирует, что буфер не отображается до завершения рисования приложения.
- Если было указано текущее целевое время, то текущее системное относительное время, которое мы ожидаем, сможет отобразить текущее текущее время, соответствующее запрошенное целевое время, примененное к настоящему приложению.


Когда система решит найти подарок для отображения на экране, он выберет последний подарок, который стал готов к показу. Если есть несколько готовых презентаций, все, кроме последних (т. е. настоящее с наибольшим имеющимся идентификатором), будут пропущены и сразу же войдете в отставленное состояние, в то время как его буфер доступных событий будет сигнализировать, но текущее отставное ограждение не будет сигнализироваться, так как пропущенное представление не переходит от
Когда будет выбрано готовое представление, система начинает работу, чтобы отобразить ее на экране. Это может означать отрисовку буфера в составе кадра DWM, а затем запрос оборудования показать этот кадр на экране, или это может означать отправку буфера непосредственно на оборудование сканирования в случае скольжения. После этого, как говорят, настоящее находится в очереди. На высоком уровне это означает, что он находится на пути к отображению.
Когда оборудование приближается к отображению настоящего, что присутствует, как сообщается, будет отображаться. Там он будет оставаться, видимым на экране, пока последующий подарок не появится и заменит его.
Когда последующий подарок становится в очереди, то мы знаем, что оборудование в конечном итоге перестанет отображать текущее текущее. На этом этапе, как говорят, настоящее уйти в отставку.
Когда этот последующий подарок отображается, то текущий подарок, как говорят, будет снят с учета.
Диспетчер презентаций предоставляет настоящий забор с выходом на пенсию, который сигнализирует настоящему идентификатору каждого присутствующего при входе в состояние выхода из эксплуатации . Этот сигнал указывает приложению, что она стала безопасной для отрисовки в буферы, связанные с этим представлением, без повреждения предыдущего представления. Если приложение выдает работу отрисовки во время выхода из эксплуатации текущего состояния, работа отрисовки будет помещена в очередь до тех пор, пока оно не войдет в состояние прекращения работы, в какой момент она будет выполнена. Если работа отрисовки будет выдана после прекращения работы, она будет выполнена немедленно.
Если работа отрисовки будет выдана после прекращения работы, она будет выполнена немедленно.
Ниже приведена схема изменения состояния.
Схема буферов, поверхностей и представлений
Ниже приведена схема, связанная с диспетчером презентаций, буферами презентаций, поверхностями презентации, презентациями и обновлениями.
На этой схеме показан диспетчер презентаций с двумя поверхностями презентации и тремя буферами презентации, которые были выпущены двумя выпущенными презентациями— первый представленный буфер 1 в поверхности 1 и буфер 2 в поверхности 2. Вторая обновленная поверхность 2 для отображения буфера презентации 3 и не изменила привязку поверхности 1. После отображения текущего 2 surface 1 будет отображать буфер 1, а поверхность 2 будет отображать буфер 3, который можно увидеть в текущем состоянии объектов в диспетчере презентаций. Каждый присутствующий в очереди вступит в силу при обработке в системе.
Примечание
Так как present 2 не изменил буфер для поверхности 1, поверхность 1 осталась привязанной к буферу 1 из предыдущего текущего.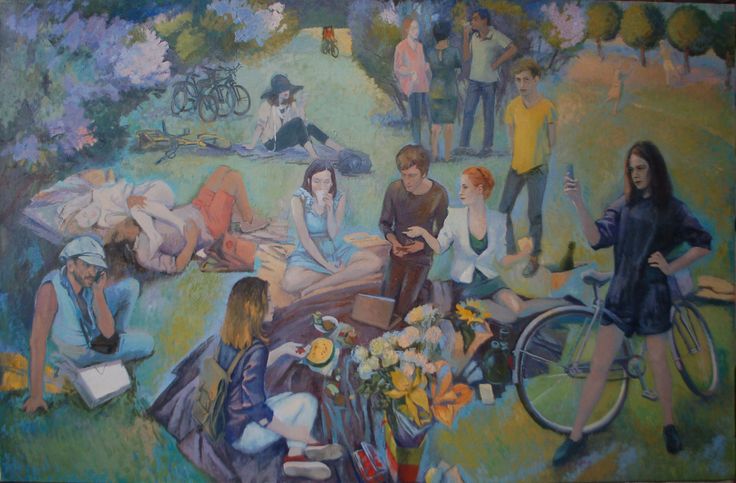 В этом смысле существует «неявная» ссылка на буфер 1 в настоящее время 2, так как поверхность 1 останется привязанной к буферу 1 после отображения 2.
В этом смысле существует «неявная» ссылка на буфер 1 в настоящее время 2, так как поверхность 1 останется привязанной к буферу 1 после отображения 2.
Добавление поверхностей презентации в визуальное дерево
Поверхности презентации — это содержимое, которое существует как часть визуального дерева композиции. Каждая поверхность презентации привязана к дескриптору поверхности композиции. В Windows. ПОЛЬЗОВАТЕЛЬСКОГО ИНТЕРФЕЙСА. Композиция, кисть поверхности может быть создана для существующего дескриптора поверхности композиции и привязана к визуальному элементу спрайта. В DirectComposition поверхность композиции может быть создана на основе существующего дескриптора поверхности композиции и привязана как содержимое к визуальному элементу. Дополнительные сведения см. в соответствующей документации по каждому API.
ТАКИЕ API, как Windows Media Foundation, созданные для использования этого API, предоставляют маркеры поверхности композиции, которые будут предварительно привязаны к поверхности презентации. Приложение также может создать собственный дескриптор поверхности композиции для последующей привязки к поверхности презентации и добавления в визуальное дерево путем вызова DCompositionCreateSurfaceHandle.
Приложение также может создать собственный дескриптор поверхности композиции для последующей привязки к поверхности презентации и добавления в визуальное дерево путем вызова DCompositionCreateSurfaceHandle.
Чтение статистики презентации
API подкачки композиции предоставляет статистику презентации, которая описывает различные сведения о том, как был обработан конкретный подарок. Как правило, информация может описать, как поверхность презентации использовалась в кадре DWM, на момент времени, в течение которого она отображалась, была ли она показана вообще, и т. д.
Существуют различные типы статистики презентаций, и они предназначены для расширения в будущих версиях API. Приложение использует диспетчер презентаций для регистрации для получения интересующих его типов статистики. Затем эти статистические данные отправляются в очередь статистики диспетчера презентаций. Диспетчер презентаций предоставляет доступное для приложений событие статистики , которое представляет собой дескриптор событий, указывающий, когда в очереди статистики есть доступные для чтения элементы статистики. Когда это делается, приложение может выключить первый элемент статистики из очереди, прочитать его и обработать его. Диспетчер презентаций сбрасывает доступное событие статистики, когда приложение считывает всю статистику в очереди. Приложение обычно считывает и обрабатывает статистику в цикле, пока не будет сброшено доступное событие статистики. Приложение обычно обрабатывает эту очередь статистики в том же рабочем цикле, который используется для выдачи. Рекомендуемый шаблон использования заключается в определении приоритета обработки статистики по сравнению с выдачей новых подарков, чтобы убедиться, что очередь не переполнена.
Когда это делается, приложение может выключить первый элемент статистики из очереди, прочитать его и обработать его. Диспетчер презентаций сбрасывает доступное событие статистики, когда приложение считывает всю статистику в очереди. Приложение обычно считывает и обрабатывает статистику в цикле, пока не будет сброшено доступное событие статистики. Приложение обычно обрабатывает эту очередь статистики в том же рабочем цикле, который используется для выдачи. Рекомендуемый шаблон использования заключается в определении приоритета обработки статистики по сравнению с выдачей новых подарков, чтобы убедиться, что очередь не переполнена.
Очередь имеет максимальное количество отслеживаемых статистических данных, которое будет находиться в порядке 512-1024 статистики. Максимальная глубина очереди должна быть достаточной, чтобы хранить статистику примерно в 5 секундах в обычных случаях. Если очередь статистики становится полной, и больше статистики сообщается, политика заключается в том, что самая старая статистика будет прекращена, чтобы освободить место.
- Цепочка буферов компоновки
- Для оптимальной производительности используйте модель флип-флип DXGI
Формируем композицию в кадре | RC.Incubator
Композиция — алгоритм творческого процесса.
Привет, уважаемые читатели! Продолжая рубрику об обучении искусству фотографии «RC.Incubator«, рассмотрим основные приемы, позволяющие нам получить эстетически красивое изображение. Более подробно об этих и других приемах вы можете узнать, придя на курс профессионального фотографа.
Что такое композиция? Композиция в фотографии представляет собой художественно-образную, содержательно-формальную целостность — наиболее сложный и совершенный тип структуры, в котором все элементы органично связаны между собой. Имеет иррациональную природу, оригинальна и неповторима. Сотворена из «простых элементов» — линия, фигура, текстура, свет, цвет, зона резкости. Несмотря на иррациональную природу композиции, она все же имеет свои правила, или скорее черты, присущие всем хорошим снимкам, используя которые в своих фотографиях, вы можете добиться высокого мастерства.
Направляющие линии
Используйте естественные линии, кривые и замкнутые контуры, вдоль которых скользит взгляд, и благодаря которым вы можете управлять взглядом зрителя. Человеческий взгляд цепляется за яркие пятна, человеческое лицо и глаза.
Использование контраста
Уместное использование контраста способно усилить и подчеркнуть красоту снимка. Это может быть контраст по яркости, цвету, форме и размеру, текстуре, масштабу.
Диагональ — это не просто отрезок
Диагональные линии это один из важных инструментов изобразительной фотографии. Благодаря им можно передать перспективу и ощущение движения на снимке, глубину кадра. Используя диагонали в кадре можно подчеркнуть статичность или динамичность сцены, указать направление движения.
1,618 золота
Классической правило «золотого сечения» и его упрощенная версия — правило третей. С этим термином мы сталкивались и за школьной партой и в стенах института. Суть правила состоит в том, что отрезок делится на две части таким образом, что длина всего отрезка относится к большей части, так же, как большая часть к меньшей. Это отношение равно числу 1.618 — золотое число, божественная пропорция. Обычно на таких силовых линиях или на их пересечении располагают значимые объекты в кадре.
Это отношение равно числу 1.618 — золотое число, божественная пропорция. Обычно на таких силовых линиях или на их пересечении располагают значимые объекты в кадре.
Симметрия и асимметрия. Баланс
В зависимости от поставленных задач в кадре важно добиваться равновесия пятен (яркостных и цветовых), а так же смысла или сознательно подчеркивать их дисбаланс. На правиле равновесия построены симметричные кадры, которые так любят перфекционисты.
Обрамление
Особенность фотографии, состоит в том, что любой снимок имеет определенные конечные размеры. Что уже само по себе создает «рамку». Для усиления акцентов можно использовать естественные обрамления внутри кадра — элементы архитектуры, ветки деревьев, причудливые очертания скал и тд.
Правило формата
Выбирайте формат кадра, подходящий к объекту и сюжету съемки. Думайте об этом до того, как нажать на кнопку спуска. В цифровой фотографии два типа форматов — прямоугольные и квадратные. Соотношение сторон у прямоугольного кадра может быть, фактически, произвольным — от стандартных 2:3, до журнальной 6:7 и к широкоформатному панорамному соотношению. Правильное использование формата подчеркнет и усилит композиционную составляющую кадра. Но никто не ограничивает вас квадратно-прямоугольным форматом — при обработке фотографий в фотошопе, вы можете кадрировать согласно вашей творческой задумке и получить кадр произвольной формы. Разумно выбранный формат, так же придаст вашему снимку дополнительную динамику или статичность.
Правильное использование формата подчеркнет и усилит композиционную составляющую кадра. Но никто не ограничивает вас квадратно-прямоугольным форматом — при обработке фотографий в фотошопе, вы можете кадрировать согласно вашей творческой задумке и получить кадр произвольной формы. Разумно выбранный формат, так же придаст вашему снимку дополнительную динамику или статичность.
Frieke Janssens
Геометрия кадра
В кадре не должно быть лишних объектов. И ключевое слово тут — СОВСЕМ. Изображенные фигуры должны быть логически связаны между собой, они могут создавать простые фигуры (треугольник, квадрат, круг).
Узор, текстура, ритм
Паттерны от англ. pattern — повторяющиеся детали, которые привносят в изображение эстетическую составляющую. Хорошо сочетаются с контрастными объектами, разбавляющими эти повторы.
Будьте проще
Простые, лаконичные кадры не менее приятны глазу, как и насыщенные динамичные. Уберите лишние объекты, упростите геометрию, подберите невыразительный фон, оставьте внимание только на одном объекте. И если вам кажется, что поймать минимализм и аскетичность в кадре легко — попробуйте:)
И если вам кажется, что поймать минимализм и аскетичность в кадре легко — попробуйте:)
Крупность плана
Хорошо когда объект съемки взаимодействует с другими составными кадра, но ни чем не хуже, когда этот объект заполняет кадр и занимает значительную его часть. Это позволит избавиться от лишнего «мусора» и волей-неволей привлечет внимание зрителя к объекту съемки.
Многоплановая композиция
В изобразительной фотографии существует три плана — передний, средний и дальний. Используется для создания объема, раскрытия идейной составляющей за счет изменения формы и размера объектов, четкости контуров.
Пространство для взгляда
Вокруг основного объекта должно быть достаточно пространства, что бы мы могли понять направление движения, проследить взглядом и не уткнуться раньше времени в край кадра.
Экспериментируйте с ракурсом!
Один и тот же объект съемки можно запечатлеть с самых неожиданных ракурсов. Не поленитесь, подумайте с какой точки съемки будет оно выглядеть наиболее выигрышно. Экспериментируйте!
Экспериментируйте!
Удачи и побольше красивых кадров!
Все фотографии взяты с сайта 1x.com , кроме снимка Frieke Janssens.
Типы композиции. Средства композиции — презентация онлайн
1. Композиция
• Термин «композиция» происходит от лат.«composition», что означает составление,
сложение, соединение частей,
приведение их в порядок.
• В энциклопедическом словаре Брокгауза
и Ефрона: «композиция – передача в
рисунке или живописи линий, форм и
образов, смутно еще рисующихся в
воображении художника, и составление
из них органичного целого, выражающего
творческий замысел художника».
2. Средства композиции
• формат,• пространство,
• композиционный центр,
• равновесие,
• ритм,
• контраст,
• светотень,
• цвет,
• декоративность,
• динамику и статику,
• симметрию и асимметрию,
• открытость и замкнутость,
• целостность.
Таким образом, средства композиции — это все, что
необходимо для ее создания, в том числе ее приемы и
правила.
 Они разнообразны, иначе их можно назвать
Они разнообразны, иначе их можно назватьсредствами художественной выразительности композиции.
Здесь названы не все, а только основные.
3. Требования композиции :
простота,ясность,
естественность,
единство,
красота.
4. Основное правило композиции
«Композициясчитается правильной,
когда ни одна ее часть
не может быть изъята
или заменена другим без
ущерба для целого»
Типы
композиции
1. замкнутая композиция
(«кулисная композиция»)
Рафаэль
«Сикстинская
мадонна»
8. 2. открытая композиция (центробежная)
Василий Поленов «Золотая осень»10. 3. симметричная композиция
Виктор Васнецов «Три богатыря»12. 4. асимметричная композиция
В. Васнецов «Витязь на распутье»14. 5. динамичная композиция
Мауриц Эшер «День и ночь»Н. Гончарова «Велосипедист»
18. 6. статичная композиция
К. Васильев«Ожидание»
20. Виды композиции
21.
 Многоплановая композиция Используется для передачи сложного исторического, жанрового или природного явления. В ней создаетсяиллюзия
Многоплановая композиция Используется для передачи сложного исторического, жанрового или природного явления. В ней создаетсяиллюзияглубины пространства. Выделяется
главная фигура, полнее раскрываются
тематические черты во всех действующих
лицах и окружающей предметной среде.
Всему изображаемому придается
наглядность и тем самым усиливается
эмоциональная реакция зрителя на
ситуацию.
23. Фризовая композиция Показывает течение жизни во времени, последовательно развивающихся событий. Все фигуры размещаются в
пределахзрительной полосы (как буквы в
строчке), они связаны друг с
другом и смыслом, и движением,
и ритмом.
(Др. Египет, Греция, иконопись).
24. Истома Савин. Владимирская Икона Божией Матери с 18 клеймами
«Загробный суд Осириса»из «Книги мертвых» Ани
26. Фронтальная композиция Двухмерность и небольшая глубина. Это панно, витражи.
29. Объемная композиция Имеет высоту, ширину, объем (глубину) и длину.
 Скульптурные изображения, архитектура.Теремной
Скульптурные изображения, архитектура.Теремнойдворец
32. Глубинно – пространственная композиция Интерьерное производство (мебель, скульптура – малая пластика, стенды и т.д.).
Оформление комнат, театров, сцен ит.д.
36. Законы композиции
37. 1. Закон целостности и неделимости композиции
38. 2. Выразительность композиции
Выразительность – это свойствохудожественного отражения
действительности.
Она проявляет себя только в законченном
произведении, являясь конечным продуктом
композиции, ее целью, вершиной.
Выразительность как воздействующее
средство композиции проявляется главным
образом в умении пользоваться контрастами.
39. Контраст
– один из художественныхприемов, в основе которого лежит
противопоставление 2 – х
соотносящихся качеств с целью их
усиления.
• Цветовые
• Светотеневые
• Основной замысел композиции может
быть построен на контрастах доброго и
злого, веселого и грустного, нового и
старого, спокойного и динамичного и т.
 п.
п.• Контраст как универсальное средство
помогает создать яркое и выразительное
произведение. Леонардо да Винчи в
«Трактате о живописи» говорил о
необходимости использовать контрасты
величин (высокого с низким, большого с
маленьким, толстого с тонким), фактур,
материалов, объема, плоскости и др.
41. 3. Уравновешенность (баланс)
— Статическая— Динамическая
Варианты уравновешивания:
•по массе и тону;
•по цвету;
•сами фигуры и пространство между
ними.
42. Динамическое уравновешивание цветом
43. Статическое уравновешивание (по оси симметрии)
44. 4. Закон равновесия
3 источника:— натура;
— закон зрительного восприятия
натуры;
— закон картинной плоскости
(формат и линия горизонта).
45. Формат
47. Линия горизонта
(линия горизонта ниже)(на уровне глаз)
(линия горизонта выше)
Осень
Весна
длинная комната
высокая комната
50. 5. Доминанта (выявление композиционного центра)
51.
 Композиционные правила
Композиционные правила52. 1. Передача движения (динамика)
Правило передачи движения:— если на картине используются одна или несколько
диагональных линий, то изображение будет казаться
более динамичным;
— эффект движения можно создать, если оставить
свободное пространство перед движущимся объектом;
— для передачи движения следует выбирать
определенный его момент, который наиболее ярко
отражает характер движения, является его
кульминацией.
54. 2. Передача покоя (статика)
Правило передачи покоя:— если на картине отсутствуют диагональные
направления;
— если перед движущимся объектом нет свободного
пространства;
— если объекты изображены в спокойных (статичных)
позах, нет кульминации действия;
— если композиция является симметричной,
уравновешенной или образует простые
геометрические схемы (треугольник, круг, овал,
квадрат, прямоугольник), то она считается
статичной.
56. 3. «Золотое сечение»
83
5
2
Образуется бесконечный ряд чисел: 1,2,3,5,8,13,21,…………
Логарифмическая кривая – «Спираль Жизни»
58.
 Выделение главного в композиции
Выделение главного в композиции59. Пространственное расположение (ближе, дальше, выше, ниже)
• Пространственное расположение(ближе, дальше, выше, ниже)
60. 2. Цвет
3. Свет4. Размер.
5. Пространство между
главной фигурой и другой
группой.
6. Движение (динамика).
62. 7. Тон
Как средство выделения силуэта63. Приемы размещения композиционного центра живописного изображения на плоскости:
Смещениекомпозиционного
центра
относительно
геометрического
центра холста
Рембрандт
«Возвращение блудного сына»
65. Доминанта
67. Смещение доминанты, относительно центра
68. Основные элементы и формы
69. Формы по происхождению: 1. Геометрические
70. Формы природные
71. Искусственно — абстрактные
Искусственно абстрактные72. Линия – основной элемент
73. Линия – элемент деления и объединения
74. Упражнения по теме «Линия»
75.
 «Жесткие линии»
«Жесткие линии»76. «Мягкие линии»
77. Упражнение по теме «Цвет»
78. Психологическая характеристика цвета
легкийтеплый
спокойный
пассивный
активный
торжественный
холодный
тяжелый
Дополнительные цвета
Упражнения
по теме «Графическая композиция»
81. 1. Метод «спирали»
2. Шрифтоваякомпозиция
Многогранные изменения в составе синапсов и вовлечение астроцитов в мышиную модель синдрома ломкой Х-хромосомы
Аннотация
Синдром ломкой Х-хромосомы (FXS), распространенная наследуемая форма умственной отсталости, как известно, изменяет неокортикальные цепи. Однако его влияние на различные типы синапсов, составляющих эти цепи, или на участие астроцитов, недостаточно известно. Мы использовали иммунофлуоресцентную матричную томографию для количественной оценки различных синаптических популяций и их связи с астроцитами в слоях с 1 по 4 соматосенсорной коры взрослых мышей модели FXS, мышей с нокаутом FMR1. Собранные многоканальные данные содержали приблизительно 1,6 миллиона синапсов, которые были проанализированы с помощью вероятностного детектора синапсов. Наше исследование выявляет сложные изменения синаптического типа и слоев в неокортикальной схеме мышей с нокаутом FMR1. Мы сообщаем об увеличении количества малых глутаматергических синапсов VGluT1 в слое 4, сопровождаемом уменьшением количества больших синапсов VGluT1 в слоях 1 и 4. Синапсы VGluT2 демонстрируют довольно последовательное снижение плотности в слоях 1 и 2/3. Во всех слоях мы наблюдаем потерю крупных тормозных синапсов. Наконец, уменьшается астроцитарная ассоциация возбуждающих синапсов. Способность анализировать дефицит цепи по типу синапса и вовлечению астроцитов будет иметь решающее значение для понимания того, как эти изменения влияют на функцию цепи, и, в конечном итоге, для определения целей для терапевтического вмешательства.
Собранные многоканальные данные содержали приблизительно 1,6 миллиона синапсов, которые были проанализированы с помощью вероятностного детектора синапсов. Наше исследование выявляет сложные изменения синаптического типа и слоев в неокортикальной схеме мышей с нокаутом FMR1. Мы сообщаем об увеличении количества малых глутаматергических синапсов VGluT1 в слое 4, сопровождаемом уменьшением количества больших синапсов VGluT1 в слоях 1 и 4. Синапсы VGluT2 демонстрируют довольно последовательное снижение плотности в слоях 1 и 2/3. Во всех слоях мы наблюдаем потерю крупных тормозных синапсов. Наконец, уменьшается астроцитарная ассоциация возбуждающих синапсов. Способность анализировать дефицит цепи по типу синапса и вовлечению астроцитов будет иметь решающее значение для понимания того, как эти изменения влияют на функцию цепи, и, в конечном итоге, для определения целей для терапевтического вмешательства.
Введение
Синдром ломкой Х-хромосомы (FXS) является наиболее распространенной наследственной формой умственной отсталости, поражающей примерно 1 из 7000 мужчин и 1 из 11000 женщин во всех расах и этнических группах 1 . Пациенты с FXS демонстрируют широкий спектр фенотипов, включая умеренную и тяжелую умственную отсталость, аутистическое поведение, макроорхизм, предрасположенность к эпилептическим припадкам и лицевые аномалии 2,3,4 . FXS вызывается молчанием гена FMR1, который кодирует белок умственной отсталости Fragile X (FMRP). Известно, что FMRP играет важную роль в трансляции, транспортировке и нацеливании большого количества мРНК в нейроны 5,6,7 . FMRP также связывается со многими белками, указывая на его участие в большом разнообразии функций, таких как регуляция стабильности генома, клеточная дифференцировка и управление ионными каналами 8. Поскольку FMRP участвует во множестве процессов в клетках, оказалось трудным понять, как дефицит FMRP влияет на синапсы и нейронные цепи в головном мозге, вызывая патологию FXS.
Пациенты с FXS демонстрируют широкий спектр фенотипов, включая умеренную и тяжелую умственную отсталость, аутистическое поведение, макроорхизм, предрасположенность к эпилептическим припадкам и лицевые аномалии 2,3,4 . FXS вызывается молчанием гена FMR1, который кодирует белок умственной отсталости Fragile X (FMRP). Известно, что FMRP играет важную роль в трансляции, транспортировке и нацеливании большого количества мРНК в нейроны 5,6,7 . FMRP также связывается со многими белками, указывая на его участие в большом разнообразии функций, таких как регуляция стабильности генома, клеточная дифференцировка и управление ионными каналами 8. Поскольку FMRP участвует во множестве процессов в клетках, оказалось трудным понять, как дефицит FMRP влияет на синапсы и нейронные цепи в головном мозге, вызывая патологию FXS.
Мышиная модель этого заболевания, мыши с нокаутом FMR1, демонстрируют фенотипы, сходные с человеческим FXS, такие как дефицит обучения и памяти 9,10,11 , сенсорная обработка 12,13 и социальное поведение 14,15 . Однако, несмотря на эти глубокие неврологические и поведенческие нарушения, сообщаемые изменения в синапсах были довольно тонкими, при этом основная синаптическая нейротрансмиссия, по-видимому, не пострадала. На синаптическом функциональном уровне мыши FMR1 KO обнаруживают специфический для региона дефицит пластичности, такой как аномальная долговременная потенциация (LTP) и длительная депрессия (LTD) 8 . Многие из молекулярных сигнальных путей в синапсах кажутся нерегулируемыми, но изменения часто специфичны для региона и типа нейрона, и вклад специфических сигнальных путей в патологию Fragile X трудно распутать 16 . На синаптическом структурном уровне наиболее очевидным отличием является более высокая плотность незрелых, длинных и тонких дендритных шипов пирамидных нейронов в коре взрослых мышей FMR1 KO по сравнению с контрольной группой WT 11,12,13,14,15 .
Однако, несмотря на эти глубокие неврологические и поведенческие нарушения, сообщаемые изменения в синапсах были довольно тонкими, при этом основная синаптическая нейротрансмиссия, по-видимому, не пострадала. На синаптическом функциональном уровне мыши FMR1 KO обнаруживают специфический для региона дефицит пластичности, такой как аномальная долговременная потенциация (LTP) и длительная депрессия (LTD) 8 . Многие из молекулярных сигнальных путей в синапсах кажутся нерегулируемыми, но изменения часто специфичны для региона и типа нейрона, и вклад специфических сигнальных путей в патологию Fragile X трудно распутать 16 . На синаптическом структурном уровне наиболее очевидным отличием является более высокая плотность незрелых, длинных и тонких дендритных шипов пирамидных нейронов в коре взрослых мышей FMR1 KO по сравнению с контрольной группой WT 11,12,13,14,15 .
Необычно длинные и тонкие шипы, которые также обнаруживаются в фиксированных тканях пациентов с FXS 9,10,17,18 , похожи на незрелые шипы, наблюдаемые во время развития 19,20,21 . Это наблюдение привело к популярной гипотезе о том, что отсутствие FMRP в нервной системе вызывает дефект созревания и обрезки позвоночника, что, в свою очередь, изменяет синаптическую связность и в конечном итоге приводит к поведенческим дефектам 6,7,22,23,24 . В то время как морфология дендритных шипов и структурная динамика являются хорошими индикаторами модификаций в синаптической связности 25,26,27 , они не могут полностью отражать разнообразие кортикальных синапсов. Например, большинство тормозных синапсов оканчиваются на дендритных стержнях и соматах и, таким образом, не объясняются изменениями шипиков. Среди возбуждающих синапсов, оканчивающихся на шипиках, выделяют кортико-кортикальные синапсы, содержащие везикулярный транспортер глутамата VGluT1, и таламокортикальные синапсы, содержащие VGluT2 28,29 , которые имеют очень разные функции в корковых схемах. Воздействие FXS, вероятно, зависит от типа синапса из-за дифференциальной экспрессии FMRP в разных типах нейронов -30-.
Это наблюдение привело к популярной гипотезе о том, что отсутствие FMRP в нервной системе вызывает дефект созревания и обрезки позвоночника, что, в свою очередь, изменяет синаптическую связность и в конечном итоге приводит к поведенческим дефектам 6,7,22,23,24 . В то время как морфология дендритных шипов и структурная динамика являются хорошими индикаторами модификаций в синаптической связности 25,26,27 , они не могут полностью отражать разнообразие кортикальных синапсов. Например, большинство тормозных синапсов оканчиваются на дендритных стержнях и соматах и, таким образом, не объясняются изменениями шипиков. Среди возбуждающих синапсов, оканчивающихся на шипиках, выделяют кортико-кортикальные синапсы, содержащие везикулярный транспортер глутамата VGluT1, и таламокортикальные синапсы, содержащие VGluT2 28,29 , которые имеют очень разные функции в корковых схемах. Воздействие FXS, вероятно, зависит от типа синапса из-за дифференциальной экспрессии FMRP в разных типах нейронов -30-. В самом деле, недавнее исследование с использованием высокомультиплексной матричной томографии показало различное влияние FXS на синаптические популяции коркового слоя 4 и 5 у мышей FMR1 KO 31 .
В самом деле, недавнее исследование с использованием высокомультиплексной матричной томографии показало различное влияние FXS на синаптические популяции коркового слоя 4 и 5 у мышей FMR1 KO 31 .
Чтобы добавить еще один уровень сложности, FXS может также воздействовать на определенные ненейронные клетки. Являясь наиболее распространенными глиальными клетками в мозге млекопитающих, астроциты модулируют синаптическую структуру и функцию 32 и связаны со многими заболеваниями нервной системы 33 . В мозге мышей астроциты также экспрессируют FMRP 34 , а мыши FMR1 KO имеют меньше синапсов гиппокампа, связанных с астроцитами 35 . Интересно, что специфическая для астроцитов делеция FMR1 приводит к значительно большему количеству незрелых шипов в моторной коре мыши из-за перепроизводства шипов во время развития 36 . Пока неизвестно, зависит ли такой вклад астроцитов от типа синапса.
Чтобы лучше понять специфические для типа синапса эффекты FXS на неокортикальную синаптическую схему, мы исследовали изменения в различных синаптических популяциях и их связь с астроцитами в соматосенсорной коре взрослых мышей, области, в которой сообщалось о различных нарушениях. для мышей FMR1 KO 37,38,39 . Мы сосредоточились на поверхностных слоях коры, где исследования с визуализацией в реальном времени выявили изменения в формировании и обороте дендритных шипиков 37,40 , но специфичность типа синапса эффектов FXS неизвестна. Чтобы исследовать большое количество синапсов разных типов, мы использовали томографию с иммунофлуоресцентной матрицей (IF-AT), которая позволяет обнаруживать уровень освещенности отдельных синапсов в ткани мозга и применять несколько маркеров для различения типов синапсов 41, 42 . Синаптическая плотность была количественно определена с использованием автоматических методов обнаружения синапсов, ранее разработанных нашей группой 43,44 . Наши результаты показывают многогранные изменения в составе и участии астроцитов в синаптических схемах соматосенсорной коры взрослых мышей FMR1 KO.
для мышей FMR1 KO 37,38,39 . Мы сосредоточились на поверхностных слоях коры, где исследования с визуализацией в реальном времени выявили изменения в формировании и обороте дендритных шипиков 37,40 , но специфичность типа синапса эффектов FXS неизвестна. Чтобы исследовать большое количество синапсов разных типов, мы использовали томографию с иммунофлуоресцентной матрицей (IF-AT), которая позволяет обнаруживать уровень освещенности отдельных синапсов в ткани мозга и применять несколько маркеров для различения типов синапсов 41, 42 . Синаптическая плотность была количественно определена с использованием автоматических методов обнаружения синапсов, ранее разработанных нашей группой 43,44 . Наши результаты показывают многогранные изменения в составе и участии астроцитов в синаптических схемах соматосенсорной коры взрослых мышей FMR1 KO.
Методы
Обзор
Раздел методов разделен на два основных компонента — создание данных и вычислительный анализ. В разделе генерации данных указаны типы используемых мышей, используемые антитела и методология визуализации. В разделе вычислительного анализа освещаются методы, используемые для автоматического анализа данных массивной томографии, включая обнаружение синапсов по их определенному типу и обнаружение астроцитов. Обработка и визуализация каждой мыши занимала в среднем десять дней, в результате чего было получено около 40 ГБ данных изображения. Сценарии анализа, запущенные параллельно, заняли одиннадцать дней на двенадцатиядерном компьютере и сгенерировали около 800 ГБ вспомогательных данных.
В разделе генерации данных указаны типы используемых мышей, используемые антитела и методология визуализации. В разделе вычислительного анализа освещаются методы, используемые для автоматического анализа данных массивной томографии, включая обнаружение синапсов по их определенному типу и обнаружение астроцитов. Обработка и визуализация каждой мыши занимала в среднем десять дней, в результате чего было получено около 40 ГБ данных изображения. Сценарии анализа, запущенные параллельно, заняли одиннадцать дней на двенадцатиядерном компьютере и сгенерировали около 800 ГБ вспомогательных данных.
Генерация данных
Исследуемые наборы данных были получены из соматосенсорной коры взрослых мышей и представляют слои с первого по четвертый. Соматосенсорная кора была выбрана из-за хорошо задокументированного дефицита у мышей FMR1 KO в этой области коры 37,38,39 . Мы сосредоточились на поверхностных слоях коры, для которых доступно больше информации посредством исследований изображений в реальном времени 37,40 . Средний объем набора данных составил 135 588 мкм 3 .
Средний объем набора данных составил 135 588 мкм 3 .
Животные
Мыши содержались группами в животноводческом комплексе Калифорнийского университета в Санта-Круз (UCSC) с 12-часовыми циклами свет-темнота и доступом к пище и воде вволю . Все процедуры проводились в соответствии с протоколами, одобренными Комитетом по уходу и использованию животных (IACUC) UCSC. Мыши FMR1 KO были получены от доктора Стивена Т. Уоррена, Университет Эмори. Мыши Thy1-YFP-H были приобретены у JAX. Всех мышей подвергали обратному скрещиванию с мышами C57BL/6 более чем в 10 поколениях для получения конгенных линий. Для текущих экспериментов самцов YFP+ WT скрещивали с самками YFP-FMR1+/-, и для экспериментов использовали только однопометных потомков самцов. Мыши WT относятся к FMR1+/y, а мыши KO относятся к FMR1-/y. Поскольку экспрессия YFP сильно различалась у разных животных, мы не использовали ее в анализе. Для анализа использовали четырех мышей KO и трех мышей WT. Мышам было четыре месяца, когда их принесли в жертву. Дополнительные сведения о мышах приведены в дополнительной таблице S1.
Мышам было четыре месяца, когда их принесли в жертву. Дополнительные сведения о мышах приведены в дополнительной таблице S1.
Матричная томография
Ткань была подготовлена с использованием стандартных протоколов матричной томографии 42 . Мышей анестезировали ингаляцией галотана, и их мозг быстро извлекали, разрезали на 2 мм среза, фиксировали погружением в 4% параформальдегид в фосфатно-солевом буфере (PBS) на 1 час при комнатной температуре, затем оставляли в фиксаторе на ночь при 4 o С. После промывки в PBS вырезали соматосенсорную кору, гасили в 50 мМ глицина в PBS в течение 30 минут и обезвожены в серии промывок этанолом (50%, 70%, 70%) при 4 900–90°С, затем пропитаны и помещены в смолу LRWhite в желатиновых капсулах и полимеризованы при 50 900–90°С. С в течение 24 часов.
Для изготовления лент серийных секций блоки обрезали вокруг ткани до формы трапеции и тонкой малярной кистью нанесли клей (Weldwood Contact Cement, разбавленный ксилолом) на переднюю и заднюю кромки пирамиды блока . Заложенный пластиковый блок разрезали на ультрамикротоме (Leica Ultracut EM UC6) на 70×9 мм.0083 нм толщиной серийных срезов, которые помещали на покровные стекла, покрытые желатином.
Заложенный пластиковый блок разрезали на ультрамикротоме (Leica Ultracut EM UC6) на 70×9 мм.0083 нм толщиной серийных срезов, которые помещали на покровные стекла, покрытые желатином.
Иммуномаркировка
Срезы обрабатывали для стандартной непрямой иммунофлуоресценции, как описано в 42 . Антитела были получены из коммерческих источников и перечислены в таблице 1. Специфические контроли для томографии представлены в дополнительной таблице S2. Срезы инкубировали в 50 мМ глицина в TBS в течение 5 минут, а затем в блокирующем растворе (0,05% Tween-20 и 0,1% BSA в TBS) в течение 5 минут. Первичные антитела разводили в блокирующем растворе, как указано в таблице 1, и наносили на 2 часа при комнатной температуре или на ночь при 4–9°С.0009 o °C. После 15-минутной промывки в TBS срезы инкубировали с вторичными антителами, конъюгированными с красителем Alexa, с высокой перекрестной адсорбцией (Life Technologies), разведенными 1:150 в блокирующем растворе, в течение 30 минут при комнатной температуре. Наконец, срезы промывали TBS в течение 15 минут, промывали дистиллированной водой и помещали на предметные стекла с использованием SlowFade Gold Antifade Mountant с DAPI (Invitrogen). После визуализации срезов антитела элюировали раствором 0,2 М NaOH и 0,02% SDS в течение 20 минут и повторно наносили новые антитела. Было применено несколько раундов элюирования и повторного окрашивания для создания высокоразмерного иммунофлуоресцентного изображения. Образцы подвергали иммуноокрашиванию бок о бок парами, состоящими из одного образца WT и одного образца KO, и визуализировали сразу после завершения окрашивания.
Наконец, срезы промывали TBS в течение 15 минут, промывали дистиллированной водой и помещали на предметные стекла с использованием SlowFade Gold Antifade Mountant с DAPI (Invitrogen). После визуализации срезов антитела элюировали раствором 0,2 М NaOH и 0,02% SDS в течение 20 минут и повторно наносили новые антитела. Было применено несколько раундов элюирования и повторного окрашивания для создания высокоразмерного иммунофлуоресцентного изображения. Образцы подвергали иммуноокрашиванию бок о бок парами, состоящими из одного образца WT и одного образца KO, и визуализировали сразу после завершения окрашивания.
Полноразмерный стол
Метод визуализации
Иммуноокрашенные ленты срезов визуализировали на автоматизированном эпифлуоресцентном микроскопе (Zeiss AxioImager Z1) с использованием масляного объектива Plan-Apochromat 1,4 NA с увеличением 63x. Для каждого участка ленты была отображена область 140 мкм × 400 мкм , и эта область охватывала слои с 1 по 4. Анализировался весь отображаемый объем, но он был разделен на отдельные подобъемы, соответствующие различным слоям. Чтобы определить список позиций для автоматического создания изображений, был использован специальный графический пользовательский интерфейс на основе Python, MosaicPlanner (полученный с https://code.google.com/archive/p/smithlabsoftware/), для автоматического поиска соответствующих местоположений по всей сети. серийные разделы. Изображения из разных сеансов визуализации были зарегистрированы с использованием красителя DAPI, присутствующего в монтажной среде. Изображения серийных срезов также выравнивали с помощью сигнала DAPI. Регистрация и выравнивание изображений выполнялись с помощью плагина MultiStackReg в FIJI 9.0009 45 .
Анализировался весь отображаемый объем, но он был разделен на отдельные подобъемы, соответствующие различным слоям. Чтобы определить список позиций для автоматического создания изображений, был использован специальный графический пользовательский интерфейс на основе Python, MosaicPlanner (полученный с https://code.google.com/archive/p/smithlabsoftware/), для автоматического поиска соответствующих местоположений по всей сети. серийные разделы. Изображения из разных сеансов визуализации были зарегистрированы с использованием красителя DAPI, присутствующего в монтажной среде. Изображения серийных срезов также выравнивали с помощью сигнала DAPI. Регистрация и выравнивание изображений выполнялись с помощью плагина MultiStackReg в FIJI 9.0009 45 .
Вычислительный анализ
Основной целью этого анализа является изучение влияния отсутствия белка FMRP на синаптический состав соматосенсорной коры. Это требует возможности количественной оценки синапсов по их молекулярному составу и их близости к астроцитарному отростку. Для этого мы взяли существующие методы и расширили их возможности для решения вычислительных задач, поставленных в этих экспериментах, включая разработку метода обнаружения отростков астроцитов, прилегающих к синапсам. Вычислительный анализ проводился вслепую. Образцы были визуализированы парами одной мыши WT и одной мыши KO с помощью MMP, а файлы обезличенных изображений были отправлены в AKS для анализа.
Для этого мы взяли существующие методы и расширили их возможности для решения вычислительных задач, поставленных в этих экспериментах, включая разработку метода обнаружения отростков астроцитов, прилегающих к синапсам. Вычислительный анализ проводился вслепую. Образцы были визуализированы парами одной мыши WT и одной мыши KO с помощью MMP, а файлы обезличенных изображений были отправлены в AKS для анализа.
Обнаружение синапсов
Для настоящих целей мы определяем «тип синапса» как определенную комбинацию синаптических белков. Например, ГАМКергический (тормозной) тип синапса определяется наличием общего пресинаптического маркера, синапсин; постсинаптический маркер тормозных синапсов — гефирин; и пресинаптический маркер тормозных синапсов, GAD. Глутаматергический (возбуждающий) синапс определяется наличием общего пресинаптического маркера, синапсин, и постсинаптический маркер для возбуждающих синапсов, PSD-95. Глутаматергический синапс с VGluT2 и рядом с астроцитарным отростком определяется наличием синапсинов, VGluT2, PSD-95 и GS, маркера астроцитов.
Обнаружение синапсов по их молекулярному составу — это первый этап вычислительного конвейера. Для количественного анализа объемов томографии большого массива жизненно важно найти подходящий метод обнаружения синапсов. Большинство опубликованных методов обнаружения синапсов используют традиционные подходы машинного обучения 43 , 46,47,48 . Все эти подходы состоят из нескольких общих шагов для обнаружения синапсов. Во-первых, для каждого типа синапсов большое количество синапсов идентифицируется вручную и помечается в массиве данных томографии. Затем классификатор (такой как машина опорных векторов или сверточная нейронная сеть) обучается с помощью этих ручных аннотаций. Наконец, весь набор данных соответствующим образом разделяется, а потенциальные синапсы помечаются классификатором. Хотя этот метод хорошо работает для некоторых вопросов биологии синапсов, сложность ручной маркировки различных типов синапсов в иммунофлуоресцентных данных делает его неэффективным для наших приложений.
Метод вероятностного обнаружения синапсов, представленный в 43 , представляет собой подход, ориентированный на тип синапса, который не требует никаких обучающих данных, что делает его жизнеспособным вариантом для исследования синапсов, полученных с помощью матричной томографии. Фокусировка на «типе синапса» означает, что пользователь указывает молекулярный состав и относительное пространственное расположение вместе с размером синаптических маркеров перед запуском метода вероятностного обнаружения синапсов. Комбинация определяемого пользователем типа синапса и размера маркера называется «запросом», как показано на рис. 1.
Рисунок 1 Схема запроса. На рисунке слева показано относительное пространственное расположение различных флуоресцентных маркеров, используемых для обнаружения возбуждающего синапса, экспрессирующего VGluT1, рядом с отростком астроцита. Это визуальное описание синапса преобразуется в запрос, показанный справа от большой черной стрелки. Запрос — это определяемое пользователем описание того, как должен «выглядеть» интересующий тип синапса. При этом ожидается, что пресинаптические белковые маркеры — синапсин и VGluT1 будут колокализоваться (занимать одно и то же трехмерное пространство) друг с другом. Кроме того, ожидается, что пресинаптические, постсинаптические и астроцитарные маркеры (как группа) будут находиться рядом друг с другом. В верхней правой части рисунка показаны три 1,5 мкм × 1,5 мкм вырезки различных комбинаций маркеров, показывающих, как выглядит запрос в данных. На первом вырезе показаны синапсин и пунктум PSD-95, наложенные друг на друга; вторая вырезка включает пунктум GS, а третий вырез включает пунктум VGluT1.
Запрос — это определяемое пользователем описание того, как должен «выглядеть» интересующий тип синапса. При этом ожидается, что пресинаптические белковые маркеры — синапсин и VGluT1 будут колокализоваться (занимать одно и то же трехмерное пространство) друг с другом. Кроме того, ожидается, что пресинаптические, постсинаптические и астроцитарные маркеры (как группа) будут находиться рядом друг с другом. В верхней правой части рисунка показаны три 1,5 мкм × 1,5 мкм вырезки различных комбинаций маркеров, показывающих, как выглядит запрос в данных. На первом вырезе показаны синапсин и пунктум PSD-95, наложенные друг на друга; вторая вырезка включает пунктум GS, а третий вырез включает пунктум VGluT1.
Полноразмерное изображение
Запрос для обнаружения глутаматергического синапса будет выглядеть следующим образом: точка PSD-95 минимум 2 пикселей × 2 пс × 2 среза (что составляет × 2 среза 0083 мкм × 0,2 мкм × 0,14 мкм для наших данных) рядом с точечным синапсином того же размера. Смежность в этом случае означает, что точки двух разных маркеров антител не занимают одно и то же пространство, а вместо этого сопоставляются друг с другом, т. е. занимают непосредственно соседние пиксели или имеют небольшую долю пикселей. Поскольку PSD-95 является постсинаптическим белком, а синапсин является пресинаптическим белком, этот простой запрос глутаматергических синапсов соответствует известной биологической модели глутаматергических синапсов. В 43 , запрос включает только пресинаптических и постсинаптических производителей. В этой работе мы расширили запрос, включив в него пресинаптические, постсинаптические и астроцитарные маркеры. Эту комбинацию маркеров часто называют «трехсторонним синапсом» 46 . Как показано в левой части рис. 1, модель трехчастного синапса предполагает, что молекулярные маркеры для каждого «подкласса» (пресинаптический, постсинаптический, астроцитарный) расположены рядом друг с другом.
Смежность в этом случае означает, что точки двух разных маркеров антител не занимают одно и то же пространство, а вместо этого сопоставляются друг с другом, т. е. занимают непосредственно соседние пиксели или имеют небольшую долю пикселей. Поскольку PSD-95 является постсинаптическим белком, а синапсин является пресинаптическим белком, этот простой запрос глутаматергических синапсов соответствует известной биологической модели глутаматергических синапсов. В 43 , запрос включает только пресинаптических и постсинаптических производителей. В этой работе мы расширили запрос, включив в него пресинаптические, постсинаптические и астроцитарные маркеры. Эту комбинацию маркеров часто называют «трехсторонним синапсом» 46 . Как показано в левой части рис. 1, модель трехчастного синапса предполагает, что молекулярные маркеры для каждого «подкласса» (пресинаптический, постсинаптический, астроцитарный) расположены рядом друг с другом.
Таким образом, вероятностный детектор синапсов — это метод обнаружения определенных типов синапсов. Вместо того, чтобы требовать, чтобы пользователь вручную аннотировал несколько экземпляров типа синапса для обучения классификатора машинного обучения, подход на основе запросов просит пользователя определить синапс, указав основные характеристики, то есть необходимые маркеры, необходимый объем точки для каждого маркер (что зависит от известного разрешения микроскопа) и их относительное пространственное расположение.
Вместо того, чтобы требовать, чтобы пользователь вручную аннотировал несколько экземпляров типа синапса для обучения классификатора машинного обучения, подход на основе запросов просит пользователя определить синапс, указав основные характеристики, то есть необходимые маркеры, необходимый объем точки для каждого маркер (что зависит от известного разрешения микроскопа) и их относительное пространственное расположение.
После создания запроса метод вероятностного обнаружения синапсов следует за запросом, чтобы автоматически обнаруживать синапсы, соответствующие запросу. На рис. 2 показан пример конвейера, идущего от необработанных входных данных к обнаружению синапсов с пороговым значением. Вкратце, синапсы обнаруживаются в данных IF-AT с помощью ряда шагов. Во-первых, пользователь определяет, как должно выглядеть обнаружение. Например, он должен состоять из точки синапсин и точки PSD-95, которые лежат рядом друг с другом, и каждая точка должна иметь размер не менее двух пикселей на два пикселя на два среза. Метод обнаружения синапсов использует эту информацию для вычисления выходного вероятностного изображения. Значение в каждом пикселе — это вероятность того, что он принадлежит определению синапса. Затем эти выходные изображения обрабатываются порогом, что приводит к обнаружению синапсов. Детали метода обнаружения синапсов описаны в 43 и его применения для характеристики антител изучаются в 44 . Оба сообщают о обширной проверке, указывающей на то, что инструмент готов решать новые биологические вопросы в этой работе.
Метод обнаружения синапсов использует эту информацию для вычисления выходного вероятностного изображения. Значение в каждом пикселе — это вероятность того, что он принадлежит определению синапса. Затем эти выходные изображения обрабатываются порогом, что приводит к обнаружению синапсов. Детали метода обнаружения синапсов описаны в 43 и его применения для характеристики антител изучаются в 44 . Оба сообщают о обширной проверке, указывающей на то, что инструмент готов решать новые биологические вопросы в этой работе.
Конвейер вероятностного обнаружения синапсов. В первом столбце показаны необработанные данные PSD-95 и синапсин. Во втором столбце показаны выходные данные метода обнаружения синапсов, где значение в каждом пикселе — это вероятность того, что пиксель принадлежит указанному определению синапса. В третьем столбце показаны результаты пороговой обработки выходной карты вероятностей. Из нашей прошлой работы мы определили эмпирический порог 0,9. быть оптимальным значением. В четвертом столбце показаны обнаружения (белым цветом), наложенные на суперпозицию данных PSD-95 и синапсов. Для этой визуализации определением синапса было смежностью PSD-95 и синапсина-пунктума минимального размера 0,2 мкм × 0,2 мкм × 0,07 мкм , что соответствует 2 PX × 2 PX PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX . × 1 срез . Смежность определяется как две точки, занимающие непосредственно соседние пиксели или разделяющие небольшую долю пикселей.
быть оптимальным значением. В четвертом столбце показаны обнаружения (белым цветом), наложенные на суперпозицию данных PSD-95 и синапсов. Для этой визуализации определением синапса было смежностью PSD-95 и синапсина-пунктума минимального размера 0,2 мкм × 0,2 мкм × 0,07 мкм , что соответствует 2 PX × 2 PX PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX . × 1 срез . Смежность определяется как две точки, занимающие непосредственно соседние пиксели или разделяющие небольшую долю пикселей.
Полноразмерное изображение
Определения типа и размера синапса
Для анализа, представленного в этой работе, мы использовали запросы, перечисленные в Таблице 2. Синапс определенного типа определяется как имеющий соответствующие маркеры, причем все маркеры являются указанного размера. Для этого исследования нам потребовалось, чтобы маркеры охватывали один или несколько срезов, в зависимости от желаемого размера синапса, и имели минимальный размер x , y 0,2 мкм × 0,2 мкм . Единственным исключением является определение синапсов VGluT2, где маркер VGluT2 должен охватывать два или более смежных среза. Это связано со свойствами антитела VGluT2, которое в дополнение к ожидаемой надежной метке субпопуляции синапсов также дает более высокий, случайно распределенный фоновый сигнал (дополнительную информацию см. в дополнительной таблице S2).
Синапс определенного типа определяется как имеющий соответствующие маркеры, причем все маркеры являются указанного размера. Для этого исследования нам потребовалось, чтобы маркеры охватывали один или несколько срезов, в зависимости от желаемого размера синапса, и имели минимальный размер x , y 0,2 мкм × 0,2 мкм . Единственным исключением является определение синапсов VGluT2, где маркер VGluT2 должен охватывать два или более смежных среза. Это связано со свойствами антитела VGluT2, которое в дополнение к ожидаемой надежной метке субпопуляции синапсов также дает более высокий, случайно распределенный фоновый сигнал (дополнительную информацию см. в дополнительной таблице S2).
Полноразмерная таблица
Размеры обнаруженных синапсов соответствовали ожидаемому логарифмически нормальному распределению 47 (дополнительный рисунок S1). Для анализа каждый тип синапса был дополнительно разделен на три размера: малый, средний и большой. Малый синапс определяется как имеющий по крайней мере один из маркеров только на одном срезе, средний синапс определяется как имеющий по крайней мере один из маркеров на двух срезах, а остальные маркеры на двух или более срезах, а большой синапс определяется как наличие всех маркеров на трех или более срезах. Чтобы вычислить плотность синапсов малых синапсов, запрос, в котором синаптические маркеры охватывают два или более слайсов, вычитается из запроса, в котором синаптические маркеры охватывают один или более слайсов. Точно так же, чтобы вычислить плотность синапсов средних синапсов, запрос, в котором синаптические маркеры охватывают три или более срезов, вычитается из запроса, в котором синаптические маркеры охватывают два или более срезов. «Все синапсы» определенного типа определяются как имеющие синаптические маркеры, охватывающие один или несколько срезов. Необходимо сделать важное уточнение относительно этой группировки синапсов по размеру. Обнаружение маркеров синапсов основано на иммунных реакциях и, следовательно, связано с ограничениями использования антител.
Малый синапс определяется как имеющий по крайней мере один из маркеров только на одном срезе, средний синапс определяется как имеющий по крайней мере один из маркеров на двух срезах, а остальные маркеры на двух или более срезах, а большой синапс определяется как наличие всех маркеров на трех или более срезах. Чтобы вычислить плотность синапсов малых синапсов, запрос, в котором синаптические маркеры охватывают два или более слайсов, вычитается из запроса, в котором синаптические маркеры охватывают один или более слайсов. Точно так же, чтобы вычислить плотность синапсов средних синапсов, запрос, в котором синаптические маркеры охватывают три или более срезов, вычитается из запроса, в котором синаптические маркеры охватывают два или более срезов. «Все синапсы» определенного типа определяются как имеющие синаптические маркеры, охватывающие один или несколько срезов. Необходимо сделать важное уточнение относительно этой группировки синапсов по размеру. Обнаружение маркеров синапсов основано на иммунных реакциях и, следовательно, связано с ограничениями использования антител. Отсутствие иммунореактивности не равнозначно отсутствию маркера, это лишь указывает на то, что уровень маркера ниже определяемого уровня. Следовательно, небольшой синапс с синапсиновой меткой только на одном срезе не равен синапсу с пресинаптическим бутоном, заключенным в объеме одной секции 70 нм , скорее, это синапс с определяемыми уровнями синапсин только на одном участке.
Отсутствие иммунореактивности не равнозначно отсутствию маркера, это лишь указывает на то, что уровень маркера ниже определяемого уровня. Следовательно, небольшой синапс с синапсиновой меткой только на одном срезе не равен синапсу с пресинаптическим бутоном, заключенным в объеме одной секции 70 нм , скорее, это синапс с определяемыми уровнями синапсин только на одном участке.
Расчет объема
Плотность синапсов рассчитывается как количество синапсов, обнаруженных в наборе данных по объему набора данных. Объем набора данных определяется объемом нейропиля (т. е. без учета объема, занимаемого ядрами клеток и крупными кровеносными сосудами, который может значительно различаться между областями). При выборе областей для изображения мы избегали крупных кровеносных сосудов. Для расчета объема клеточных ядер ядерный краситель DAPI был преобразован в вероятностное пространство с использованием методов, описанных в 9.0009 44 . Вкратце, значение для каждого пикселя в вероятностном пространстве — это вероятность того, что он принадлежит переднему плану, в диапазоне от 0 до 1. Для этого моделируется фоновый шум, и вероятность переднего плана равна единице минус вероятность того, что пиксель принадлежит переднему плану. фон. Фоновый шум моделируется как гауссовский, для которого среднее значение и дисперсия рассчитываются из самих необработанных данных. После того, как карта вероятностей рассчитана, для нее устанавливается пороговое значение (t = 0,6, выбранное путем наблюдения) и очищается с помощью последовательности морфологических операций. Код доступен для скачивания на сайте проекта. Таким образом, объем нейропиля был получен путем вычитания объема ядер, окрашенных DAPI, из общего отображаемого объема.
Для этого моделируется фоновый шум, и вероятность переднего плана равна единице минус вероятность того, что пиксель принадлежит переднему плану. фон. Фоновый шум моделируется как гауссовский, для которого среднее значение и дисперсия рассчитываются из самих необработанных данных. После того, как карта вероятностей рассчитана, для нее устанавливается пороговое значение (t = 0,6, выбранное путем наблюдения) и очищается с помощью последовательности морфологических операций. Код доступен для скачивания на сайте проекта. Таким образом, объем нейропиля был получен путем вычитания объема ядер, окрашенных DAPI, из общего отображаемого объема.
Анализ колокализации
Для изучения пространственных отношений между глиальными и постсинаптическими маркерами мы использовали тест колокализации на Фиджи 45 , применяя метод рандомизации ван Стенселя 48 . Для каждой пары каналов мы использовали интересующую область размером 40 × 26 мкм через стек изображений. Для сравнения использовали пары возбуждающих и тормозных пресинаптических/постсинаптических маркеров, VGluT1/PSD95 и GAD/Gephyrin.
Для сравнения использовали пары возбуждающих и тормозных пресинаптических/постсинаптических маркеров, VGluT1/PSD95 и GAD/Gephyrin.
Статистический анализ
Статистический анализ для определения значимости между двумя популяциями был проведен с помощью двустороннего непарного t-критерия, который показал свою применимость к очень небольшим размерам выборки 49 , таким как использованный в этом исследовании (n = 4). Из-за потенциальных проблем с любым статистическим тестом на небольшой выборке мы также представляем данные по каждой отдельной выборке в разделе дополнительных рисунков. Графики, показывающие данные для каждого типа синапса, размера и слоя, можно найти на дополнительных рисунках S2, S3, S4, S5, S6 и S7. Альтернативный статистический анализ может быть выполнен с данными и кодом, которые мы публично предоставляем.
Результаты
Мы использовали иммунофлуоресцентную матричную томографию (IF-AT) для количественной оценки синаптической плотности, состава и участия глии в слоях с 1 по 4 соматосенсорной коры взрослых мышей с нокаутом FMR1 (KO) и мышей дикого типа (WT). . IF-AT основан на цифровой реконструкции изображений, полученных из массивов серийных ультратонких срезов (70 нм), прикрепленных к покровным стеклам, иммунофлуоресцентно помеченных и визуализированных под флуоресцентным микроскопом. Использование ультратонких срезов позволяет определять уровень освещенности отдельных синапсов, а возможность применения нескольких иммунофлуоресцентных маркеров (10 и более) позволяет идентифицировать различные синаптические популяции 41 . Синаптическая плотность была количественно определена с использованием автоматических методов обнаружения синапсов, ранее разработанных нашей группой 43,44 . Помимо уже опубликованной проверки нашего метода, результаты мышей WT также сравнивали с доступными оценками в литературе в качестве дополнительного контроля.
. IF-AT основан на цифровой реконструкции изображений, полученных из массивов серийных ультратонких срезов (70 нм), прикрепленных к покровным стеклам, иммунофлуоресцентно помеченных и визуализированных под флуоресцентным микроскопом. Использование ультратонких срезов позволяет определять уровень освещенности отдельных синапсов, а возможность применения нескольких иммунофлуоресцентных маркеров (10 и более) позволяет идентифицировать различные синаптические популяции 41 . Синаптическая плотность была количественно определена с использованием автоматических методов обнаружения синапсов, ранее разработанных нашей группой 43,44 . Помимо уже опубликованной проверки нашего метода, результаты мышей WT также сравнивали с доступными оценками в литературе в качестве дополнительного контроля.
Обзор наборов данных и обнаруженных синапсов
Объемы примерно 140 × 400 × 2,7 мкм охватывающих слоев с 1 по 4 соматосенсорной коры мышей FMR1 KO и мышей WT были визуализированы, как показано на рис. 3. Мы обнаружили в среднем 200 000 синапсов в каждом томе, всего около 1,6 миллиона синапсов во всех восьми наборах данных. Возбуждающие синапсы идентифицировали по наличию иммунофлуоресцентных сигналов как от синапсинов, пресинаптических белков, так и от PSD-9.5 — белок постсинаптического каркаса возбуждающих синапсов. Возбуждающие синапсы далее подразделялись в зависимости от их везикулярных переносчиков глутамата на VGluT1-положительные, обычно считающиеся внутрикорковыми, и VGluT2-положительные, принадлежащие преимущественно к таламокортикальным входам 28,29,50 . Тормозные синапсы идентифицировали по наличию общего пресинаптического маркера синапсин и пресинаптического маркера ГАМКергических синапсов, декарбоксилазы глутаминовой кислоты (GAD), вместе с постсинаптическим маркером гефирином. Астроциты, включая их отростки, выявляли с помощью антител против глутаминсинтетазы (ГС) 51,52 , что позволило выделить долю синапсов, непосредственно примыкающих к астроцитарным отросткам.
3. Мы обнаружили в среднем 200 000 синапсов в каждом томе, всего около 1,6 миллиона синапсов во всех восьми наборах данных. Возбуждающие синапсы идентифицировали по наличию иммунофлуоресцентных сигналов как от синапсинов, пресинаптических белков, так и от PSD-9.5 — белок постсинаптического каркаса возбуждающих синапсов. Возбуждающие синапсы далее подразделялись в зависимости от их везикулярных переносчиков глутамата на VGluT1-положительные, обычно считающиеся внутрикорковыми, и VGluT2-положительные, принадлежащие преимущественно к таламокортикальным входам 28,29,50 . Тормозные синапсы идентифицировали по наличию общего пресинаптического маркера синапсин и пресинаптического маркера ГАМКергических синапсов, декарбоксилазы глутаминовой кислоты (GAD), вместе с постсинаптическим маркером гефирином. Астроциты, включая их отростки, выявляли с помощью антител против глутаминсинтетазы (ГС) 51,52 , что позволило выделить долю синапсов, непосредственно примыкающих к астроцитарным отросткам. В дополнение к идентификации синапсов на основе комбинаций различных маркеров синапсы также анализировались на основе их размера, поскольку известно, что размер коррелирует со зрелостью и силой синаптической связи. Вновь образованные синапсы имеют тенденцию быть небольшими, а в зрелых синапсах известно, что размер постсинаптической плотности хорошо коррелирует с синаптической силой 53 . Визуальный осмотр наборов данных не выявил каких-либо очевидных различий в интенсивности и характере иммунофлуоресценции для любого из маркеров между мышами KO и WT. Толщина коры также была сопоставима между двумя условиями (0,88 ± 0,03 мм для KO против 0,89 ± 0,02 мм для WT, p = 0,71).
В дополнение к идентификации синапсов на основе комбинаций различных маркеров синапсы также анализировались на основе их размера, поскольку известно, что размер коррелирует со зрелостью и силой синаптической связи. Вновь образованные синапсы имеют тенденцию быть небольшими, а в зрелых синапсах известно, что размер постсинаптической плотности хорошо коррелирует с синаптической силой 53 . Визуальный осмотр наборов данных не выявил каких-либо очевидных различий в интенсивности и характере иммунофлуоресценции для любого из маркеров между мышами KO и WT. Толщина коры также была сопоставима между двумя условиями (0,88 ± 0,03 мм для KO против 0,89 ± 0,02 мм для WT, p = 0,71).
Обзор наборов данных. ( A ) Иммунофлуоресцентная матричная томография соматосенсорной коры мышей дикого типа и мышей с нокаутом FMR1. На левой панели показана область изображения в каждом образце, состоящая из четырех плиток, охватывающих слои коры с 1 по 4. Окрашивание ядер DAPI, объемная реконструкция 30 серийных срезов по 70 нм каждый. На правой панели показана иммунофлуоресценция синаптических (синапсин, VGluT1, VGluT2 и GAD2) и глиальных (GS) маркеров в слое 4 соматосенсорной коры мышей дикого типа и нокаутных мышей, объемная реконструкция 10 серийных срезов, 70 нм каждый. ( B ) Сводка изменений плотности пунктата в одном канале между мышами дикого типа и мышами с нокаутом с усреднением всех слоев. Графики показывают процентную разницу между мышами дикого типа и мышами с нокаутом и имеют цветовую кодировку, чтобы указать направление изменения и статистическую значимость. ( C ) На этом графике показано распределение плотности точек GS по размеру. «Маленькие» точки охватывают один срез, «средние» точки охватывают два среза, а «большие» точки охватывают три или более срезов.
Окрашивание ядер DAPI, объемная реконструкция 30 серийных срезов по 70 нм каждый. На правой панели показана иммунофлуоресценция синаптических (синапсин, VGluT1, VGluT2 и GAD2) и глиальных (GS) маркеров в слое 4 соматосенсорной коры мышей дикого типа и нокаутных мышей, объемная реконструкция 10 серийных срезов, 70 нм каждый. ( B ) Сводка изменений плотности пунктата в одном канале между мышами дикого типа и мышами с нокаутом с усреднением всех слоев. Графики показывают процентную разницу между мышами дикого типа и мышами с нокаутом и имеют цветовую кодировку, чтобы указать направление изменения и статистическую значимость. ( C ) На этом графике показано распределение плотности точек GS по размеру. «Маленькие» точки охватывают один срез, «средние» точки охватывают два среза, а «большие» точки охватывают три или более срезов.
Полноразмерное изображение
Одноканальный анализ
Сначала мы количественно оценили плотность точек различных синаптических маркеров у мышей KO и WT. Это довольно грубый анализ, особенно для небольших точек, которые присутствуют только на одном срезе и поэтому потенциально могут включать неспецифическую метку. Однако эти результаты дают обзор данных, а также подчеркивают важность последующего обнаружения синапсов как гораздо более надежного инструмента для мониторинга синаптических изменений. Количественная оценка плотности точек различных синаптических маркеров не выявила каких-либо статистически значимых различий (9).0083 p > 0,05) при усреднении слоев 1–4, как показано на рис. 3B и более подробно на дополнительном рис. S2. Отмечена тенденция к уменьшению количества мелких точек VGluT2 ( p = 0,065) и крупных точек PSD95 ( p = 0,065). С другой стороны, средние точки PSD-95 имели тенденцию к увеличению ( p = 0,056), как показано на средней панели рис. 3. размеры: у мышей FMR1 KO было больше мелких точек ( p = 0,01), а также тенденция к уменьшению количества крупных точек ЗС ( p = 0,07), как показано на нижней панели рис.
Это довольно грубый анализ, особенно для небольших точек, которые присутствуют только на одном срезе и поэтому потенциально могут включать неспецифическую метку. Однако эти результаты дают обзор данных, а также подчеркивают важность последующего обнаружения синапсов как гораздо более надежного инструмента для мониторинга синаптических изменений. Количественная оценка плотности точек различных синаптических маркеров не выявила каких-либо статистически значимых различий (9).0083 p > 0,05) при усреднении слоев 1–4, как показано на рис. 3B и более подробно на дополнительном рис. S2. Отмечена тенденция к уменьшению количества мелких точек VGluT2 ( p = 0,065) и крупных точек PSD95 ( p = 0,065). С другой стороны, средние точки PSD-95 имели тенденцию к увеличению ( p = 0,056), как показано на средней панели рис. 3. размеры: у мышей FMR1 KO было больше мелких точек ( p = 0,01), а также тенденция к уменьшению количества крупных точек ЗС ( p = 0,07), как показано на нижней панели рис. 3.
3.
значительные изменения в плотности точек различных синаптических маркеров, это не обязательно исключает изменения в синаптических популяциях мышей FMR1 KO. Синаптические белки, оцененные в нашем исследовании, действительно сильно обогащены в синапсах, но они также обнаружены в экстрасинаптических участках, и, таким образом, их присутствие не обязательно означает наличие синапса.
Общая плотность синапсов
Гораздо более точное обнаружение синапсов достигается при использовании комбинаций генераторов синапсов, в идеале по крайней мере одного пресинаптического и одного постсинаптического маркера, как указано в нашем алгоритме обнаружения синапсов 43,44 . Действительно, с использованием таких комбинаций синаптических маркеров обнаруженные плотности и распределения синапсов у мышей дикого типа согласуются с предыдущими оценками, как показано на рис. 4. Известно, что подавляющее большинство корковых синапсов являются либо возбуждающими глутаматергическими, либо тормозящими ГАМКергическими синапсами 54 . Таким образом, общую плотность синапсов оценивали по сумме плотностей обнаруженных глутаматергических (синапсин + маркеры PSD95) и ГАМКергических синапсов (синапсин + GAD + гефириновые маркеры), что дает примерно 1,94 синапса на мкм 3 встроенных салфетка. Поскольку обезвоживание ткани и встраивание в соответствии с нашим протоколом приводят к линейной усадке примерно на 23% или объемной усадке на 54% 55 , это соответствует 0,9 синапсам на мкм 3 необработанной ткани, что очень похоже на зарегистрированную плотность синапсов в коре головного мозга мыши 56 .
Таким образом, общую плотность синапсов оценивали по сумме плотностей обнаруженных глутаматергических (синапсин + маркеры PSD95) и ГАМКергических синапсов (синапсин + GAD + гефириновые маркеры), что дает примерно 1,94 синапса на мкм 3 встроенных салфетка. Поскольку обезвоживание ткани и встраивание в соответствии с нашим протоколом приводят к линейной усадке примерно на 23% или объемной усадке на 54% 55 , это соответствует 0,9 синапсам на мкм 3 необработанной ткани, что очень похоже на зарегистрированную плотность синапсов в коре головного мозга мыши 56 .
Обзор распределения плотности синапсов дикого типа. ( A ) Вверху, мультиплексная иммунофлуоресценция из той же области одного среза соматосенсорной коры мыши дикого типа. Ядерное пятно DAPI окрашено в голубой цвет. Белыми квадратами выделены отдельные синапсы, которые на А внизу показаны с большим увеличением на 4 последовательных последовательных срезах. ( B ) Распределение типов синапсов по разным слоям. Слои со значительными различиями для типа синапса отмечены. Хотя нет существенной разницы в плотности слоев для глутаматергических синапсов в целом и для глутаматергических синапсов с VGluT1, существует значительная разница между слоями для глутаматергических синапсов с VGluT2, как с VGluT1, так и с VGluT2, а также для тормозных синапсов. ( C ) Фракция тормозных синапсов в слоях с 1 по 4 соматосенсорной коры мыши. ( D ) Фракция синапсов VGluT2 в слоях с 1 по 4 соматосенсорной коры мыши.
( B ) Распределение типов синапсов по разным слоям. Слои со значительными различиями для типа синапса отмечены. Хотя нет существенной разницы в плотности слоев для глутаматергических синапсов в целом и для глутаматергических синапсов с VGluT1, существует значительная разница между слоями для глутаматергических синапсов с VGluT2, как с VGluT1, так и с VGluT2, а также для тормозных синапсов. ( C ) Фракция тормозных синапсов в слоях с 1 по 4 соматосенсорной коры мыши. ( D ) Фракция синапсов VGluT2 в слоях с 1 по 4 соматосенсорной коры мыши.
Изображение полного размера
Относительные вклады тормозных и возбуждающих типов синапсов, которые мы обнаруживаем, согласуются с известным составом синапсов коры мыши. Процент тормозных ГАМКергических синапсов в наших обнаружениях варьируется от 12% в слое 1 до 20% в слое 4 (рис. 4F). Электронно-микроскопические (ЭМ) подсчеты в соматосенсорной коре мыши показывают, что тормозные синапсы состоят из 11% синапсов в слое 1-9. 0009 57 и 18% синапсов в слое 4 58 .
0009 57 и 18% синапсов в слое 4 58 .
Распределение синапсов VGluT2 по слоям согласуется с их известным предпочтением слоя 4 (рис. 4G). Известно, что VGluT2 метит таламокортикальные синапсы, которые нацелены в основном на слой 4 и нижний слой 2/3 59,60 . Было показано, что таламокортикальные синапсы, идентифицированные либо методами дегенерации, антероградным транспортом лектина, либо иммуноокрашиванием VGluT2, составляют примерно 20% глутаматергических синапсов в слое 4 соматосенсорной коры мыши 61,62,63 и мы действительно видим 21% в слое 4.
Для дальнейшей проверки точности наших обнаружений мы использовали другой способ расчета плотности возбуждающих глутаматергических синапсов. Эти синапсы можно разделить на две основные популяции в зависимости от везикулярных переносчиков глутамата (VGluT) в пресинаптическом участке, при этом большинство возбуждающих синапсов содержат VGluT1, и меньшую популяцию, в основном сосредоточенную в слое 4, VGluT2. Кроме того, некоторые синапсы экспрессируют как VGluT1, так и VGluT2 9.0009 64 . Таким образом, плотность глутаматергических синапсов должна быть примерно равна плотности синапсов VGluT1 и VGluT2 за вычетом синапсов VGluT1/VGluT2, чтобы предотвратить двойной подсчет одного и того же синапса. Это действительно имело место, как видно из Таблицы 3.
Кроме того, некоторые синапсы экспрессируют как VGluT1, так и VGluT2 9.0009 64 . Таким образом, плотность глутаматергических синапсов должна быть примерно равна плотности синапсов VGluT1 и VGluT2 за вычетом синапсов VGluT1/VGluT2, чтобы предотвратить двойной подсчет одного и того же синапса. Это действительно имело место, как видно из Таблицы 3.
Полный размер таблицы
Изменения в плотности синапсов у мышей FMR1 KO
Затем мы сравнили плотности различных популяций синапсов у мышей WT и мышей FMR1 KO. Несмотря на то, что отдельные точки синаптических маркеров не показали каких-либо статистически значимых различий в двух состояниях, были широкомасштабные изменения в плотности синапсов. Эти изменения зависели от типа и размера синапса, а также от коркового слоя, как показано на рис. 5 и более подробно на дополнительном рис. S3. Наблюдалось увеличение количества малых глутаматергических синапсов VGluT1 в слое 4, сопровождавшееся уменьшением количества больших синапсов VGluT1 в слоях 1 и 4. С другой стороны, синапсы, содержащие VGluT2, демонстрировали довольно последовательное снижение плотности в слоях 1 и 2/3. . Крупные тормозные синапсы уменьшились во всех исследованных слоях, при этом не было выявлено изменений в малых и средних тормозных синапсах.
С другой стороны, синапсы, содержащие VGluT2, демонстрировали довольно последовательное снижение плотности в слоях 1 и 2/3. . Крупные тормозные синапсы уменьшились во всех исследованных слоях, при этом не было выявлено изменений в малых и средних тормозных синапсах.
Изменения плотности синапсов по слоям. ( A ) Сводная таблица, показывающая типы синапсов, которые имеют значительное увеличение или уменьшение плотности синапсов между мышами дикого типа и мышами с нокаутом. В каждой строке показаны графики процентной разницы в плотности между мышами дикого типа и мышами с нокаутом. Графики имеют цветовую кодировку, чтобы указать направление изменения и статистическую значимость. ( B – D ) Различия в плотности синапсов для разных типов синапсов для трех ожидаемых размеров синапсов.
Изображение в натуральную величину
Эти изменения в плотности различных синаптических популяций привели к увеличению соотношения возбуждение-торможение у мышей FMR1 KO, что согласуется с предыдущими отчетами: 6,20 ± 0,56 для KO против 5,24 ± 0,32 у мышей WT. (среднее для слоев с 1 по 4; p = 0,04). Аналогичное увеличение наблюдалось во всех исследованных слоях, но оно не достигало статистической значимости. Возбуждение определяли по количеству синапсов, содержащих синапсин и PSD95, а торможение — по количеству синапсов, содержащих синапс, GAD и гефирин. Эти изменения выделены на рис. 6G.
(среднее для слоев с 1 по 4; p = 0,04). Аналогичное увеличение наблюдалось во всех исследованных слоях, но оно не достигало статистической значимости. Возбуждение определяли по количеству синапсов, содержащих синапсин и PSD95, а торможение — по количеству синапсов, содержащих синапс, GAD и гефирин. Эти изменения выделены на рис. 6G.
Распределение размеров постсинаптических маркеров. ( A ) Сводная таблица, показывающая различия в распределении размеров точек PSD-95, связанных с синапсами по типу. Основной вывод заключается в том, что для возбуждающих синапсов в целом наблюдается увеличение количества маленьких точек PSD-95 у мышей с нокаутом FMR1, в то время как количество очень больших точек PSD-95, связанных с синапсами, содержащими VGluT2, уменьшается. ( B – E ) Графики, показывающие распределение PSD-95 точек как для мышей дикого типа, так и для мышей с нокаутом. ( F ) Распределение размеров гефириновых точек, связанных с тормозными синапсами. ( G ) График показывает значительное увеличение соотношения возбуждающих и тормозных синапсов у мышей с нокаутом.
( G ) График показывает значительное увеличение соотношения возбуждающих и тормозных синапсов у мышей с нокаутом.
Изображение в натуральную величину
Поскольку известно, что сила возбуждающих синапсов пропорциональна размеру постсинаптической плотности, мы также проанализировали изменения конкретно на постсинаптической стороне, как показано на рис. 6A–F и более подробно в Дополнении. Рис. S4. Выявлено статистически значимое увеличение плотности глутаматергических и конкретно VGluT1-синапсов при малых размерах PSD-9.5 точек (охватывающих только 1 или 2 среза) и значительное снижение плотности синапсов VGluT2 с большими точками PSD-95 (охватывающими 6 или более срезов). Никаких изменений плотности тормозных синапсов в зависимости от размера гефириновых точек обнаружено не было, как более подробно показано на дополнительном рисунке S5.
Вовлечение глии (астроцитов)
Астроциты тесно вовлечены в синаптическую функцию, и их отростки обнаруживаются рядом со многими синаптическими щелями в неокортексе. Поскольку астроциты мыши также экспрессируют FMRP 34 и предположительно играют роль в патогенезе FXS 36 , мы проанализировали потенциальные изменения в вовлечении астроцитов в синапсы у мышей FMR1 KO. Астроциты были обнаружены с использованием антител к глутаминсинтетазе, ферменту, который, как известно, экспрессируется преимущественно этим типом клеток 51,52 и особенно обнаружен в периферических астроцитарных отростках, которые контактируют с синапсами 65,66 . На рисунке 7A показан пример данных.
Поскольку астроциты мыши также экспрессируют FMRP 34 и предположительно играют роль в патогенезе FXS 36 , мы проанализировали потенциальные изменения в вовлечении астроцитов в синапсы у мышей FMR1 KO. Астроциты были обнаружены с использованием антител к глутаминсинтетазе, ферменту, который, как известно, экспрессируется преимущественно этим типом клеток 51,52 и особенно обнаружен в периферических астроцитарных отростках, которые контактируют с синапсами 65,66 . На рисунке 7A показан пример данных.
Астроцитарная ассоциация синапсов у мышей дикого типа. ( A ) Часть данных о синапсе дикого типа, где синапс выделен белым прямоугольником. Ниже приведены последовательные срезы выделенного синапса, показанные при большем увеличении. ( B ) Доля синапсов, связанных с астроцитами, варьируется в зависимости от типа синапса. ( C ) Показатель взаимной корреляции как функция бокового смещения между двумя каналами. Для сравнения показаны пары синаптических маркеров VGluT1 — PSD-95 и GAD — Gephyrin. Корреляция между синаптическими маркерами высока без сдвига и резко падает при латеральном смещении, как и ожидалось для тесно коррелированных пресинаптических и постсинаптических маркеров. С другой стороны, GAD и VGluT1 не совместно локализуются, потому что они маркируют разные типы синапсов, и показатель отрицательной колокализации без сдвига постепенно увеличивается до 0 при латеральном смещении. ( D ) Межслойные различия в астроцитарной ассоциации разных типов синапсов.
Для сравнения показаны пары синаптических маркеров VGluT1 — PSD-95 и GAD — Gephyrin. Корреляция между синаптическими маркерами высока без сдвига и резко падает при латеральном смещении, как и ожидалось для тесно коррелированных пресинаптических и постсинаптических маркеров. С другой стороны, GAD и VGluT1 не совместно локализуются, потому что они маркируют разные типы синапсов, и показатель отрицательной колокализации без сдвига постепенно увеличивается до 0 при латеральном смещении. ( D ) Межслойные различия в астроцитарной ассоциации разных типов синапсов.
Увеличенное изображение
У мышей дикого типа мы обнаружили, что большинство глутаматергических синапсов (72 ± 2%) примыкают к астроцитарным отросткам, что было обнаружено с помощью иммунометки к глутаминсинтетазе. Это очень похоже на предыдущие оценки ЭМ в слое 4 соматосенсорной коры мышей, где 68% глутаматергических синапсов на дендритных шипах контактировали с астроцитарными отростками на границе бутон-шип 67 . Доля глутаматергических синапсов, контактирующих с астроцитами, не была одинаковой по слоям, и мы обнаружили самую высокую ассоциацию в слое 2/3 (рис. 7D), как и в предыдущих наблюдениях в зрительной коре крыс 68 . Интересно, что мы наблюдали значительные различия в астроцитарной ассоциации различных типов синапсов, как показано на рис. 7B. Таким образом, по сравнению с большинством глутаматергических синапсов, содержащих VGluT1 (61 ± 2% астроцитарной ассоциации), значительно меньше синапсов VGluT2 (46 ± 1%, p < 0,001) прилегали к астроцитам. Разница в астроцитарной ассоциации была еще более выраженной при рассмотрении тормозных синапсов GAD, которые в два раза реже примыкали к астроцитарному отростку (29 ± 1%, p < 0,001) по сравнению с возбуждающими синапсами VGluT1.
Доля глутаматергических синапсов, контактирующих с астроцитами, не была одинаковой по слоям, и мы обнаружили самую высокую ассоциацию в слое 2/3 (рис. 7D), как и в предыдущих наблюдениях в зрительной коре крыс 68 . Интересно, что мы наблюдали значительные различия в астроцитарной ассоциации различных типов синапсов, как показано на рис. 7B. Таким образом, по сравнению с большинством глутаматергических синапсов, содержащих VGluT1 (61 ± 2% астроцитарной ассоциации), значительно меньше синапсов VGluT2 (46 ± 1%, p < 0,001) прилегали к астроцитам. Разница в астроцитарной ассоциации была еще более выраженной при рассмотрении тормозных синапсов GAD, которые в два раза реже примыкали к астроцитарному отростку (29 ± 1%, p < 0,001) по сравнению с возбуждающими синапсами VGluT1.
Поскольку ранее не было данных о различиях в глиальной ассоциации кортикальных возбуждающих и тормозных синапсов, мы искали дополнительные доказательства, чтобы подтвердить этот вывод. Мы количественно определили колокализацию между астроцитарным маркером (GS) и постсинаптическими маркерами возбуждения (PSD9).5) и тормозные (гефириновые) синапсы по методу ван Стенселя 48 . Этот метод оценивает степень пространственной корреляции путем проверки влияния очень небольших относительных смещений между парами изображений маркеров на измерение перекрытия изображений, коэффициент корреляции Пирсона. Из-за обилия синаптических и глиальных маркеров перекрывающиеся пространственные распределения могут возникать случайно. Однако если связь между двумя каналами реальна, то любое смещение одного канала относительно другого уменьшит наблюдаемую степень колокализации. С другой стороны, если два канала имеют тенденцию быть взаимоисключающими, то сдвиг увеличит степень колокализации. Наконец, если ассоциация между двумя каналами происходит случайно, то сдвиг существенно не повлияет на степень колокализации. Поскольку этот анализ проводился на объемах, содержащих несколько тысяч синапсов, которые в коре имеют случайную ориентацию, изменение степени колокализации между двумя каналами зависит не от направления сдвига, а только от его размера.
Мы количественно определили колокализацию между астроцитарным маркером (GS) и постсинаптическими маркерами возбуждения (PSD9).5) и тормозные (гефириновые) синапсы по методу ван Стенселя 48 . Этот метод оценивает степень пространственной корреляции путем проверки влияния очень небольших относительных смещений между парами изображений маркеров на измерение перекрытия изображений, коэффициент корреляции Пирсона. Из-за обилия синаптических и глиальных маркеров перекрывающиеся пространственные распределения могут возникать случайно. Однако если связь между двумя каналами реальна, то любое смещение одного канала относительно другого уменьшит наблюдаемую степень колокализации. С другой стороны, если два канала имеют тенденцию быть взаимоисключающими, то сдвиг увеличит степень колокализации. Наконец, если ассоциация между двумя каналами происходит случайно, то сдвиг существенно не повлияет на степень колокализации. Поскольку этот анализ проводился на объемах, содержащих несколько тысяч синапсов, которые в коре имеют случайную ориентацию, изменение степени колокализации между двумя каналами зависит не от направления сдвига, а только от его размера. В соответствии с нашими выводами с помощью вероятностного детектора синапсов метод ван Стенселя обнаружил колокализацию между астроцитарным маркером GS и постсинаптическим возбуждающим маркером PSD9.5 (рис. 7C). Степень колокализации увеличивалась со сдвигом между двумя каналами до 0,3–0,4 мкм , что позволяет предположить, что два маркера являются соседними, но не перекрываются. Колокализации между GS и маркером постсинаптического торможения гефирином обнаружено не было. Это не означает, что GS не присутствовал рядом с частью тормозных синапсов, это означает только то, что GS не был преимущественно связан с тормозными синапсами. Этот результат согласуется с нашим выводом о том, что возбуждающие синапсы с большей вероятностью связаны с астроцитарными процессами по сравнению с тормозными синапсами.
В соответствии с нашими выводами с помощью вероятностного детектора синапсов метод ван Стенселя обнаружил колокализацию между астроцитарным маркером GS и постсинаптическим возбуждающим маркером PSD9.5 (рис. 7C). Степень колокализации увеличивалась со сдвигом между двумя каналами до 0,3–0,4 мкм , что позволяет предположить, что два маркера являются соседними, но не перекрываются. Колокализации между GS и маркером постсинаптического торможения гефирином обнаружено не было. Это не означает, что GS не присутствовал рядом с частью тормозных синапсов, это означает только то, что GS не был преимущественно связан с тормозными синапсами. Этот результат согласуется с нашим выводом о том, что возбуждающие синапсы с большей вероятностью связаны с астроцитарными процессами по сравнению с тормозными синапсами.
Сравнение мышей дикого типа с нокаутированными мышами выявило ряд существенных изменений в вовлечении астроцитов в синапсы. В соответствии с обнаруженными общими изменениями в плотности синапсов наблюдалось значительное снижение плотности синапсов, прилегающих к астроцитам, почти для всех типов и размеров синапсов, как показано на рис. 8A и более подробно на дополнительном рис. S6. Единственным исключением были небольшие глутаматергические синапсы в слое 4, для которых плотность синапсов, прилегающих к астроцитам, увеличивалась у нокаутированных мышей. В то время как на плотность синапсов, прилегающих к астроцитам, очень сильно влияют изменения общей синаптической плотности, доля синапсов, прилегающих к астроцитам, отражает фактические изменения в вовлечении глии при синдроме ломкой Х-хромосомы. Отмечено общее снижение доли синапсов, прилегающих к астроцитам, для всех типов и слоев синапсов, однако во многих случаях различия не достигали статистической значимости, вероятно, из-за большой вариабельности внутри каждого генотипа. Значительное уменьшение доли синапсов, прилегающих к астроцитам, было обнаружено для глутаматергических синапсов и синапсов VGluT1 всех размеров, за исключением небольших синапсов VGluT1 (объединенные слои 1–4), как показано на рис. 8B и более подробно на дополнительном рисунке S7. При анализе по слоям коры наблюдалось значительное снижение доли астроцитарной ассоциации для больших глутаматергических синапсов в слоях 1 и 2/3, больших синапсов VGluT1 в слое 1, малых и средних синапсов VGluT2 в слоях 2/3 и средних синапсов GAD.
8A и более подробно на дополнительном рис. S6. Единственным исключением были небольшие глутаматергические синапсы в слое 4, для которых плотность синапсов, прилегающих к астроцитам, увеличивалась у нокаутированных мышей. В то время как на плотность синапсов, прилегающих к астроцитам, очень сильно влияют изменения общей синаптической плотности, доля синапсов, прилегающих к астроцитам, отражает фактические изменения в вовлечении глии при синдроме ломкой Х-хромосомы. Отмечено общее снижение доли синапсов, прилегающих к астроцитам, для всех типов и слоев синапсов, однако во многих случаях различия не достигали статистической значимости, вероятно, из-за большой вариабельности внутри каждого генотипа. Значительное уменьшение доли синапсов, прилегающих к астроцитам, было обнаружено для глутаматергических синапсов и синапсов VGluT1 всех размеров, за исключением небольших синапсов VGluT1 (объединенные слои 1–4), как показано на рис. 8B и более подробно на дополнительном рисунке S7. При анализе по слоям коры наблюдалось значительное снижение доли астроцитарной ассоциации для больших глутаматергических синапсов в слоях 1 и 2/3, больших синапсов VGluT1 в слое 1, малых и средних синапсов VGluT2 в слоях 2/3 и средних синапсов GAD. в слое 1.
в слое 1.
Резюме различий плотности синапсов астроцитов у нокаутных мышей. ( A ) Изменения показаны как процентная разница в плотности между мышами дикого типа и мышами с нокаутом. Графики имеют цветовую кодировку, чтобы указать направление изменения и статистическую значимость. Плотность уменьшается между мышами дикого типа и нокаутными мышами для слоев 1–3, и разница особенно заметна для средних и больших синапсов. ( B ) Изменения доли синапсов, прилегающих к астроцитарным отросткам.
Изображение в натуральную величину
Обсуждение
Используя иммунофлуоресцентную матричную томографию и методы автоматического вероятностного обнаружения синапсов, мы показали широкомасштабные изменения синапсов и их связь с астроцитами в соматосенсорной коре взрослых мышей с нокаутом FMR1, модель Fragile X. . В целом отмечается значительное снижение плотности возбуждающих глутаматергических синапсов и их связи с астроцитами. Однако изменения сильно различаются, а иногда и в противоположных направлениях, в зависимости от типа синапса, размера, а также коркового слоя. Изменения в надгранулярных слоях (слои 1 и 2/3) отражают общее снижение плотности глутаматергических синапсов, преимущественно типа VGluT2. В то же время в зернистом слое (слой 4) наблюдается небольшое, но значительное увеличение плотности глутаматергических синапсов в результате увеличения малых синапсов VGluT1, компенсируемое уменьшением больших синапсов VGluT1. Уменьшается астроцитарная ассоциация возбуждающих синапсов. Что касается тормозных ГАМКергических синапсов, то наблюдается последовательное снижение плотности крупных синапсов во всех исследованных слоях и уменьшение астроцитарной ассоциации средних синапсов в слое 1. Общая потеря тормозных синапсов больше, чем возбуждающих. синапсов, что свидетельствует о потенциальном сдвиге соотношения возбуждения/торможения в сторону большего возбуждения у мышей с нокаутом FMR1. Т.о., отсутствие FMRP заметно изменяет неокортикальную синаптическую схему как за счет изменения относительного вклада синапсов разных типов, так и за счет вовлечения астроцитов в синапсы.
Однако изменения сильно различаются, а иногда и в противоположных направлениях, в зависимости от типа синапса, размера, а также коркового слоя. Изменения в надгранулярных слоях (слои 1 и 2/3) отражают общее снижение плотности глутаматергических синапсов, преимущественно типа VGluT2. В то же время в зернистом слое (слой 4) наблюдается небольшое, но значительное увеличение плотности глутаматергических синапсов в результате увеличения малых синапсов VGluT1, компенсируемое уменьшением больших синапсов VGluT1. Уменьшается астроцитарная ассоциация возбуждающих синапсов. Что касается тормозных ГАМКергических синапсов, то наблюдается последовательное снижение плотности крупных синапсов во всех исследованных слоях и уменьшение астроцитарной ассоциации средних синапсов в слое 1. Общая потеря тормозных синапсов больше, чем возбуждающих. синапсов, что свидетельствует о потенциальном сдвиге соотношения возбуждения/торможения в сторону большего возбуждения у мышей с нокаутом FMR1. Т.о., отсутствие FMRP заметно изменяет неокортикальную синаптическую схему как за счет изменения относительного вклада синапсов разных типов, так и за счет вовлечения астроцитов в синапсы.
Наши результаты согласуются с опубликованными данными о взрослых мышах с нокаутом FMR1, предполагающих изменения кортикальной цепи, такие как увеличение незрелых синапсов меньшего размера и уменьшение синапсов большего размера (о чем свидетельствует размер шипа в 35,69 ), как а также снижение ассоциации астроцитов с синапсами гиппокампа 35 . Мы расширили эти наблюдения, показав, что эти изменения неоднородны, а зависят от типа синапса, а также коркового слоя. Предыдущее исследование действительно показало, что синапсы слоев 4 и 5 разных типов в соматосенсорной коре мыши демонстрируют различные дефициты у мышей с нокаутом FMR1 31 , и теперь мы также охарактеризовали специфические изменения синаптического типа в надгранулярных слоях. Наконец, мы впервые показываем уменьшение вовлечения астроцитов в возбуждающие глутаматергические синапсы неокортекса, наиболее выраженное в синапсах среднего и большого размера.
Возможным недостатком нашего исследования является небольшое количество проанализированных животных из-за трудоемкости этих экспериментов. Чтобы свести к минимуму различия между экспериментальными сеансами и получить более точные результаты, наш первоначальный дизайн заключался в том, чтобы иммуномаркировать и отображать образцы парами, один KO и один WT. Последующий анализ показал хорошую согласованность от эксперимента к эксперименту, о чем свидетельствуют отдельные точки данных, представленные на дополнительных рисунках, что позволило обнаружить множественные синаптические изменения у мышей FMR1 KO независимо от небольшого количества животных. Кроме того, сравнение наших результатов на мышах WT с доступными опубликованными данными еще раз подтвердило, что мы не только можем правильно определить плотность двух основных типов синапсов, возбуждающих и тормозных, но также обнаруживаем известные вариации слоев. в синапсах VGluT2 61,62,63 , и даже довольно тонкие вариации слоя в астроцитарной ассоциации возбуждающих синапсов 68 . Тем не менее, довольно большая изменчивость внутри генотипа для некоторых типов синапсов и слоев коры, вероятно, не позволила нам обнаружить все изменения в синапсах мышей FMR1 KO.
Чтобы свести к минимуму различия между экспериментальными сеансами и получить более точные результаты, наш первоначальный дизайн заключался в том, чтобы иммуномаркировать и отображать образцы парами, один KO и один WT. Последующий анализ показал хорошую согласованность от эксперимента к эксперименту, о чем свидетельствуют отдельные точки данных, представленные на дополнительных рисунках, что позволило обнаружить множественные синаптические изменения у мышей FMR1 KO независимо от небольшого количества животных. Кроме того, сравнение наших результатов на мышах WT с доступными опубликованными данными еще раз подтвердило, что мы не только можем правильно определить плотность двух основных типов синапсов, возбуждающих и тормозных, но также обнаруживаем известные вариации слоев. в синапсах VGluT2 61,62,63 , и даже довольно тонкие вариации слоя в астроцитарной ассоциации возбуждающих синапсов 68 . Тем не менее, довольно большая изменчивость внутри генотипа для некоторых типов синапсов и слоев коры, вероятно, не позволила нам обнаружить все изменения в синапсах мышей FMR1 KO. Таким образом, тот факт, что мы не наблюдали статистически значимых изменений в вовлечении астроцитов в синапсы в слое 4, не следует интерпретировать как свидетельство отсутствия изменений. Для окончательного решения этого вопроса необходимы дальнейшие эксперименты с участием большего количества животных.
Таким образом, тот факт, что мы не наблюдали статистически значимых изменений в вовлечении астроцитов в синапсы в слое 4, не следует интерпретировать как свидетельство отсутствия изменений. Для окончательного решения этого вопроса необходимы дальнейшие эксперименты с участием большего количества животных.
Наши результаты поднимают важные вопросы о последствиях наблюдаемых изменений в неокортикальных синаптических схемах мышей FMR1 KO. Эти изменения сложны и включают различные типы синапсов, а также глию. Наблюдается сдвиг размеров возбуждающих, предположительно внутрикортикальных VGluT1-синапсов, проявляющийся в увеличении малых VGluT1-синапсов и уменьшении крупных VGluT1-синапсов. Поскольку известно, что размер синапса коррелирует с силой синапса, это изменение подразумевает уменьшение более сильных синапсов VGluT1. Также наблюдается снижение плотности VGluT2, предположительно таламокортикальных синапсов, также известных как сильные синапсы 70 . Одновременно наблюдается уменьшение участия астроцитов в глутаматергических синапсах, что, вероятно, приводит к нарушению захвата глутамата 71 . Комбинация этих изменений предполагает снижение передачи глутаматергических сигналов в соматосенсорной коре мышей FMR1 KO, возникающее из-за меньшего количества сильных внутрикортикальных и таламокортикальных синапсов. Это может сопровождаться потерей остроты этих связей, поскольку меньшее количество тесно связанных астроцитов может привести к большему выбросу глутамата в эти синапсы. Сопутствующая потеря тормозных синапсов может в некоторой степени противодействовать потере силы возбуждающего сигнала, но также может способствовать потере остроты зрения, существенно снижая функциональное отношение сигнал/шум цепи у мышей FMR1 KO. Известно, что соматосенсорная обработка нарушена у мышей FMR1 KO 12,13 , а снижение отношения сигнал/шум может способствовать многим дефицитам, например, размытым картам усов, наблюдаемым у мышей FMR1 KO 72,73 . Интересно, что снижение сенсорной настройки наблюдается в слоях 2/3, но не в слое 4 соматосенсорной коры FMR1 KO мышей 72 , что согласуется с гораздо большими изменениями в синаптических схемах слоя 2/3, о которых мы сообщаем.
Комбинация этих изменений предполагает снижение передачи глутаматергических сигналов в соматосенсорной коре мышей FMR1 KO, возникающее из-за меньшего количества сильных внутрикортикальных и таламокортикальных синапсов. Это может сопровождаться потерей остроты этих связей, поскольку меньшее количество тесно связанных астроцитов может привести к большему выбросу глутамата в эти синапсы. Сопутствующая потеря тормозных синапсов может в некоторой степени противодействовать потере силы возбуждающего сигнала, но также может способствовать потере остроты зрения, существенно снижая функциональное отношение сигнал/шум цепи у мышей FMR1 KO. Известно, что соматосенсорная обработка нарушена у мышей FMR1 KO 12,13 , а снижение отношения сигнал/шум может способствовать многим дефицитам, например, размытым картам усов, наблюдаемым у мышей FMR1 KO 72,73 . Интересно, что снижение сенсорной настройки наблюдается в слоях 2/3, но не в слое 4 соматосенсорной коры FMR1 KO мышей 72 , что согласуется с гораздо большими изменениями в синаптических схемах слоя 2/3, о которых мы сообщаем.
Распространенная гипотеза функционального дефицита при аутизме фокусируется на дисбалансе возбуждения и торможения 74 . Хотя наши результаты согласуются с увеличением отношения возбуждения/торможения, они также подчеркивают сложность изменений, которые могут способствовать этому эффекту, и поддерживают представление о том, что отношение возбуждения/торможения является мерой, объединяющей различные изменения в организме. синаптические схемы, которые, вероятно, имеют различные функциональные последствия 75 . Кроме того, мы также показываем, что, помимо синапсов, существуют специфические дефициты кортикальных астроцитов и их взаимодействия с синапсами. Наиболее выраженные изменения происходят в глутаматергических синапсах. Действительно, было показано, что глутаматергические, но не ГАМКергические нейроны критически зависят от присутствия глии для установления синаптической передачи 76 . Исследования, использующие селективную делецию FMRP в астроцитах, убедительно свидетельствуют об участии этих глиальных клеток в патогенезе Fragile X , 36, , вероятно, из-за нарушения захвата глутамата , 71, . Интересно, что уменьшение астроцитарной ассоциации глутаматергических синапсов может по-разному влиять на синапсы VGluT1 и VGluT2 и, таким образом, может способствовать гетерогенным изменениям в этих двух типах синапсов. Таким образом, было показано, что при развитии зрительной коры астроциты секретируют гевин, белок, который специфически стабилизирует синапсы VGluT2 9.0009 77 . Уменьшение присутствия астроцитов в глутаматергических синапсах и, следовательно, снижение концентрации гевина может привести к потере некоторых синапсов VGluT2 без влияния на синапсы VGluT1. Таким образом, оказывается, что при Fragile X Syndrome астроциты могут опосредовать, по крайней мере, некоторые из патологических эффектов на глутаматергические синапсы 78,79,80 .
Интересно, что уменьшение астроцитарной ассоциации глутаматергических синапсов может по-разному влиять на синапсы VGluT1 и VGluT2 и, таким образом, может способствовать гетерогенным изменениям в этих двух типах синапсов. Таким образом, было показано, что при развитии зрительной коры астроциты секретируют гевин, белок, который специфически стабилизирует синапсы VGluT2 9.0009 77 . Уменьшение присутствия астроцитов в глутаматергических синапсах и, следовательно, снижение концентрации гевина может привести к потере некоторых синапсов VGluT2 без влияния на синапсы VGluT1. Таким образом, оказывается, что при Fragile X Syndrome астроциты могут опосредовать, по крайней мере, некоторые из патологических эффектов на глутаматергические синапсы 78,79,80 .
В целом, наше исследование выявило комплексные изменения синаптического типа и слоев в соматосенсорной коре мышей с нокаутом FMR1. Некоторые из этих изменений имеют противоположные направления или затрагивают только небольшую популяцию синапсов и поэтому становятся неясными при анализе общего синаптического содержимого. Способность анализировать дефициты по конкретным категориям синапсов, а также вовлечение астроцитов имеет решающее значение для понимания общей картины синаптических изменений, чтобы начать разгадывать многочисленные способы, которыми они влияют на функцию цепи, и, в конечном итоге, определить цели для терапевтического лечения и профилактика.
Способность анализировать дефициты по конкретным категориям синапсов, а также вовлечение астроцитов имеет решающее значение для понимания общей картины синаптических изменений, чтобы начать разгадывать многочисленные способы, которыми они влияют на функцию цепи, и, в конечном итоге, определить цели для терапевтического лечения и профилактика.
Доступность данных и кода
Код и необработанные данные доступны для загрузки по адресу https://aksimhal.github.io/astrocytes-synapses-fxs/.
Ссылки
Hunter, J. и др. . Эпидемиология синдрома ломкой Х-хромосомы: систематический обзор и метаанализ. Американский журнал медицинской генетики, часть A 164 , 1648–1658, https://doi.org/10.1002/ajmg.a.36511 (2014).
Артикул Google ученый
Берри-Кравис, Э. Эпилепсия при синдроме ломкой Х-хромосомы.
Развивающая медицина и детская неврология 44 , 724–728, https://doi.org/10.1111/j.1469-8749.2002.tb00277.x (2002).Артикул Google ученый
Фиш, Г. С., Сименсен, Р. Дж. и Шроер, Р. Дж. Продольные изменения показателей когнитивного и адаптивного поведения у детей и подростков с мутацией Fragile X или аутизмом. Journal of Autism and Developmental Disorders 32 , 107–114, https://doi.org/10.1023/A:1014888505185 (2002).
Артикул пабмед Google ученый
Пенагарикано О., Мулле Дж. Г. и Уоррен С. Т. Патофизиология синдрома ломкой Х-хромосомы. Ежегодный обзор геномики и генетики человека 8 , 109–129, https://doi.org/10.1146/annurev.genom.8.080706.0
(2007).КАС Статья пабмед Google ученый
Bagni, C. & Greenough, W. T. От трафика мРНП до дисморфогенеза позвоночника: корни синдрома ломкой Х-хромосомы. Nature Reviews Neuroscience 6 , 376–387, https://doi.org/10.1038/nrn1667 (2005).
КАС Статья пабмед Google ученый
Бардони, Б., Давидович, Л., Бенсаид, М. и Ханджян, Э. В. Синдром хрупкой Х-хромосомы: изучение его молекулярной основы и поиск лечения. Expert Reviews in Molecular Medicine 8 , 1–16, https://doi.org/10.1017/S1462399406010751 (2006).
Артикул пабмед Google ученый
Wisniewski, K.E., Segan, S.M., Miezejeski, C.M., Sersen, E.A. & Rudelli, R.D. Синдром fra(X): неврологические, электрофизиологические и нейропатологические отклонения. Американский журнал медицинской генетики 38 , 476–480, https://doi.org/10.1002/ajmg.1320380267 (1991).
КАС Статья пабмед Google ученый
Irwin, SA и др. . Аномальные характеристики дендритных шипов в височной и зрительной коре у пациентов с синдромом ломкой Х-хромосомы: количественное исследование. Американский журнал медицинской генетики 98 , 161–167, 10.
1002/1096-8628(20010115)98:2<161::AID-AJMG1025>3.0.CO;2-B (2001).КАС Статья Google ученый
Comery, T. A. и др. . Аномальные дендритные шипы у хрупких мышей с нокаутом X: дефицит созревания и обрезки. Proceedings of the National Academy of Sciences 94 , 5401–5404, https://doi.org/10.1073/pnas.94.10.5401 (1997).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
Irwin, SA и др. . Характеристики дендритного шипа и дендритного поля пирамидальных нейронов слоя V в зрительной коре мышей с нокаутом ломкой Х-хромосомы. Американский журнал медицинской генетики 111 , 140–146, https://doi.org/10.1002/ajmg.10500 (2002).
Артикул пабмед Google ученый
Galvez, R. & Greenough, W. T. Последовательность аномального развития дендритных шипов в первичной соматосенсорной коре мышиной модели синдрома ломкой X умственной отсталости.
Американский журнал медицинской генетики 135 A , 155–160, https://doi.org/10.1002/ajmg.a.30709 (2005).Артикул Google ученый
McKinney, B.C., Grossman, A.W., Elisseou, N.M. & Greenough, W.T. Аномалии дендритных шипов в затылочной коре у мышей с нокаутом C57BL/6 Fmr1. Американский журнал медицинской генетики — нейропсихиатрическая генетика 136 B , 98–102, https://doi.org/10.1002/ajmg.b.30183 (2005).
Артикул Google ученый
Grossman, A.W., Elisseou, N.M., McKinney, B.C. & Greenough, W.T. Пирамидальные клетки гиппокампа у взрослых мышей с нокаутом Fmr1 демонстрируют незрелый профиль дендритных шипиков. Brain Research 1084 , 158–164, https://doi.org/10.1016/j.brainres.2006.02.044 (2006).
КАС Статья пабмед Google ученый
Руделли, Р. Д. и др. . Синдром ломкой Х-хромосомы у взрослых — клинико-невропатологические данные. Acta Neuropathologica 67 , 289–295, https://doi.org/10.1007/BF00687814 (1985).
КАС Статья пабмед Google ученый
Хинтон В. Дж., Браун В. Т., Вишневски К. и Руделли Р. Д. Анализ неокортекса у трех мужчин с синдромом ломкой Х-хромосомы. Американский журнал медицинской генетики 41 , 289–294, https://doi.org/10.1002/ajmg.1320410306 (1991).
КАС Статья пабмед Google ученый
Фиала, Дж. К., Файнберг, М., Попов, В. и Харрис, К. М. Синаптогенез через дендритные филоподии в развивающейся области гиппокампа CA1. Журнал нейробиологии: официальный журнал Общества нейробиологов 18 , 8900–11, https://doi.org/10.1523/JNEUROSCI.18-21-08900.1998 (1998).
КАС Статья Google ученый
Zuo, Y., Lin, A., Chang, P. & Gan, W. B. Развитие долговременной стабильности дендритного позвоночника в различных областях коры головного мозга. Нейрон 46 , 181–189, https://doi.org/10.1016/j.neuron.2005.04.001 (2005).
КАС Статья пабмед Google ученый
Гриноу, В.
Т. и др. . Синаптическая регуляция синтеза белка и хрупкого белка X. Proceedings of the National Academy of Sciences 98 , 7101–7106, https://doi.org/10.1073/pnas.141145998 (2001).ОБЪЯВЛЕНИЕ КАС Статья Google ученый
Черчилль, Дж. Д. и др. . Подход конвергентных методов к синдрому ломкой Х-хромосомы. Психобиология развития 40 , 323–338, https://doi.org/10.1002/dev.10036 (2002).
Артикул пабмед Google ученый
Бюро И., Шеперд Г. М. Г. и Свобода К. Дефекты цепи и пластичности в развивающейся соматосенсорной коре мышей с нокаутом Fmr1. Journal of Neuroscience 28 , 5178–5188, https://doi.org/10.1523/JNEUROSCI.1076-08.2008, NIHMS150003 (2008).
КАС Статья Google ученый
Касаи Х., Фукуда М., Ватанабэ С., Хаяси-Такаги А. и Ногучи Дж. Структурная динамика дендритных шипиков в памяти и познании. Trends in Neurosciences 33 , 121–129, https://doi.org/10.1016/j.tins.2010.01.001, arXiv:1011.1669v3 (2010).
КАС Статья Google ученый
Сала, К. и Сигал, М. Дендритные шипы: местонахождение структурной и функциональной пластичности. Physiological Reviews 94 , 141–188, https://doi.org/10.1152/physrev.00012.2013 (2014).
КАС Статья пабмед Google ученый
Грациано А., Лю Х.-Б., Мюррей К.Д. и Джонс Э.Г. Два набора глутаматергических афферентов к соматосенсорному таламусу и две таламокортикальные проекции у мыши. J Comp Neurol 507 , 1258–76, https://doi.org/10.1002/cne.21592 (2008).
КАС Статья пабмед Google ученый
Zorio, D.A.R., Jackson, C.M., Liu, Y., Rubel, E.W. & Wang, Y. Клеточное распределение хрупкого белка умственной отсталости X в мозге мыши. Journal of Comparative Neurology 525 , 818–849, https://doi.org/10.1002/cne.24100 (2017).
КАС Статья пабмед Google ученый
Верхратский А. и Недергаард М. Астроглиальная колыбель в жизни синапса. Philosophical Transactions of the Royal Society B: Biological Sciences 369 , https://doi.org/10.1098/rstb.2013.0595 (2014).
Артикул Google ученый
Молофск А. В. и др. . Астроциты и болезни: перспектива развития нервной системы. Гены и развитие 26 , 891–907, https://doi.org/10.1101/gad.188326.112 (2012 г.).
КАС Статья Google ученый
Джаваид, С. и др. . Изменения в синапсах гиппокампа CA1 в мышиной модели синдрома ломкой Х-хромосомы. Глия 66 , 789–800, https://doi.org/10.1002/glia.23284 (2018).
Артикул пабмед Google ученый
Ходжес, Дж. Л. и др. . Вклад астроцитов в синаптические нарушения и нарушения обучения в мышиной модели синдрома ломкой Х-хромосомы. Биологическая психиатрия 82 , 139–149, https://doi.org/10.1016/j.biopsych.2016.08.036 (2017).
КАС Статья пабмед Google ученый
Гонсалвес, Дж. Т., Ансти, Дж. Э., Гольшани, П. и Портера-Кайо, К. Дефекты на уровне цепи в развивающемся неокортексе мышей Fragile X. Nature Neuroscience 16 , 903–909, https://doi.org/10.1038/nn.3415, 15334406 (2013).
Артикул Google ученый
Хе, Синтия, X. и др. . Тактильная защита и нарушение адаптации нейронной активности в модели аутизма у мышей с нокаутом Fmr1. Journal of Neuroscience 37 , 6475–6487, https://doi.org/10.
1523/JNEUROSCI.0651-17.2017 (2017).КАС Статья пабмед Google ученый
Cruz-Martin, A., Crespo, M. & Portera-Cailliau, C. Отсроченная стабилизация дендритных шипов у хрупких мышей X. Журнал неврологии 30 , 7793–7803 (2010).
КАС Статья Google ученый
Мичева, К. Д. и Смит, С. Дж. Массивная томография: новый инструмент для визуализации молекулярной архитектуры и ультраструктуры нейронных цепей. Нейрон 55 , 25–36, https://doi.org/10.1016/j.neuron.2007.06.014, arXiv:1011.1669v3 (2007).
КАС Статья Google ученый
Мичева, К. Д., Буссе, Б., Вейлер, Н. К., О’Рурк, Н. и Смит, С. Дж. Анализ одного синапса разнообразной популяции синапсов: методы протеомной визуализации и маркеры. Нейрон 68 , 639–53, https://doi.
org/10.1016/j.neuron.2010.09.024 (2010).КАС Статья пабмед ПабМед Центральный Google ученый
Симхал, А. К. и др. . Вероятностное обнаружение синапсов на основе флуоресценции. PLoS Computational Biology 13 , e1005493, https://doi.org/10.1371/journal.pcbi.1005493, 1611.05479 (2017).
ОБЪЯВЛЕНИЕ Статья Google ученый
Симхал, А. К. и др. . Инструмент для компьютерной характеристики синаптических антител для томографии. Frontiers in Neuroanatomy 12 , 258756, https://doi.org/10.3389/fnana.2018.00051 (2018).
КАС Статья Google ученый
Шинделин, Дж. и др. . Фиджи: платформа с открытым исходным кодом для анализа биологических изображений. Природные методы 9 , 676–82, https://doi.
org/10.1038/nmeth.2019 (2012).КАС Статья пабмед Google ученый
Буссе Б. и Смит С. Автоматический анализ разнообразной популяции синапсов. PLoS Вычислительная биология 9 , e1002976, https://doi.org/10.1371/journal.pcbi.1002976 (2013).
ОБЪЯВЛЕНИЕ КАС Статья пабмед ПабМед Центральный Google ученый
Крешук, А., Кете, У., Пакс, Э., Бок, Д. Д. и Хампрехт, Ф. А. Автоматическое обнаружение синапсов в стеках изображений просвечивающей электронной микроскопии серийных срезов. PLoS ONE 9 , e87351, https://doi.org/10.1371/journal.pone.0087351 (2014).
ОБЪЯВЛЕНИЕ КАС Статья пабмед ПабМед Центральный Google ученый
Коллман, Ф. и др. . Картирование синапсов методом сопряженной светоэлектронной матричной томографии.
Journal of Neuroscience 35 , 5792–5807, https://doi.org/10.1523/JNEUROSCI.4274-14.2015 (2015).КАС Статья пабмед Google ученый
Басс К., Хелккула П., Де Паола В., Клопат К. и Бхарат А. А. Обнаружение аксональных синапсов на трехмерных двухфотонных изображениях. ПЛОС ОДИН 12 , e0183309, https://doi.org/10.1371/journal.pone.0183309 (2017).
КАС Статья пабмед ПабМед Центральный Google ученый
Араке А., Парпура В., Санзгири Р. П. и Хейдон П. Г. Трехсторонние синапсы: глия, непризнанный партнер. Trends in Neurosciences 22 , 208–215 (1999).
КАС Статья Google ученый
Мерчан-Перес, А. и др. . Трехмерное пространственное распределение синапсов в неокортексе: исследование двухлучевой электронной микроскопии.
Кора головного мозга 24 , 1579–1588 (2013).Артикул Google ученый
Ван Стенсел, Б. и др. . Частичная колокализация глюкокортикоидных и минералокортикоидных рецепторов в дискретных компартментах ядер нейронов гиппокампа крысы. Journal of Cell Science 792 , 787–792 (1996).
Google ученый
де Винтер, Дж. К. Ф. Использование t-критерия Стьюдента с чрезвычайно малыми размерами выборки. Практическая оценка, исследования и оценка 18 , 1–12 (2013).
MathSciNet Google ученый
Канеко Т. и Фудзияма Ф. Комплементарное распределение везикулярных переносчиков глутамата в центральной нервной системе. Neuroscience Research 42 , 243–250, https://doi.org/10.1016/S0168-0102(02)00009-3 (2002).
КАС Статья пабмед Google ученый
Мартинес-Эрнандес, А., Белл, К.П. и Норенберг, М.Д. Глутаминсинтетаза: глиальная локализация в головном мозге. Science 195 , 1356–1358, https://doi.org/10.1126/science.14400 (1977).
ОБЪЯВЛЕНИЕ КАС Статья пабмед Google ученый
Anlauf, E. & Derouiche, A. Глутаминсинтетаза как астроцитарный маркер: ее клеточный тип и локализация пузырьков. Frontiers in Endocrinology 4 , 144, https://doi.org/10.3389/fendo.2013.00144 (2013).
Артикул пабмед ПабМед Центральный Google ученый
Холтмаат А. и Свобода К. Зависимая от опыта структурная синаптическая пластичность в мозге млекопитающих. Nature Reviews Neuroscience 10 , 647 (2009).
КАС Статья Google ученый
ДеФелипе Дж., Алонсо-Нанкларес Л. и Арельяно Дж. И. Микроструктура неокортекса: сравнительные аспекты. Journal of Neurocytology 31 , 299–316, https://doi.org/10.1023/A:1024130211265 (2002).
Артикул пабмед Google ученый
Schüz, A. & Palm, G. Плотность нейронов и синапсов. The Journal Of Comparative Neurology 455 , 442–455, https://doi.org/10.1371/journal.pone.00 (1989).
КАС Статья Google ученый
Кали, К. и др. . Влияние старения на структуру нейропиля в соматосенсорной коре мыши — анализ слоя 1 с помощью трехмерной электронной микроскопии. PLoS ONE 13 , e0198131, https://doi.org/10.1371/journal.pone.0198131 (2018).
КАС Статья пабмед ПабМед Центральный Google ученый
Коралек, К. А., Дженсен, К. Ф. и Киллаки, Х. П. Доказательства двух взаимодополняющих паттернов таламического входа в соматосенсорную кору крысы. Brain Research 463 , 346–351, https://doi.org/10.1016/0006-8993(88)-8 (1988).
КАС Статья пабмед Google ученый
Мейер Х.С. и др. . Специфическая для типа клеток таламическая иннервация в столбце вибриссальной коры крысы. Кора головного мозга 20 , 2287–2303, https://doi.org/10.1093/cercor/bhq069 (2018).
Артикул Google ученый
Келлер А., Уайт Э. Л. и Чиполлони П. Б. Идентификация окончаний таламокортикальных аксонов в стволах коры мыши Sml с использованием иммуногистохимии антероградно транспортируемого лектина (Phaseolus vulgaris-leucoagglutinin). Исследование мозга 343 , 159–165 (1985).
КАС Статья Google ученый
Бопп, Р., Холлер-Рикауэр, С., Мартин, К. А. и Шухкнехт, Г. Ф. Ультраструктурное исследование таламического входа в слой 4 первичной моторной и первичной соматосенсорной коры у мышей.
The Journal of Neuroscience 37 , 2435–2448, https://doi.org/10.1523/JNEUROSCI.2557-16.2017 (2017).КАС Статья пабмед ПабМед Центральный Google ученый
Накамура, К. и др. . Временно повышенная совместная локализация везикулярных переносчиков глутамата 1 и 2 на окончаниях одиночных аксонов во время постнатального развития неокортекса мышей: количественный анализ с коэффициентом корреляции. European Journal of Neuroscience 28 , 1032–1046, https://doi.org/10.1111/j.1460-9568.2008.06449.x (2008).
Артикул Google ученый
Reichenbach, A., Derouiche, A. & Kirchhoff, F. Морфология и динамика перисинаптической глии. Brain Research Reviews 63 , 11–25, https://doi.org/10.1016/j.brainresrev.2010.02.003 (2010).
Артикул пабмед Google ученый
Genoud, C. и др. . Пластичность астроцитарного покрытия и экспрессия переносчика глутамата в коре головного мозга взрослых мышей. PLOS Biology 4 , 1–8, https://doi.org/10.1371/journal.pbio.0040343 (2006).
КАС Статья Google ученый
Джонс, Т. А. и Гриноу, В. Т. Ультраструктурные доказательства усиленного контакта между астроцитами и синапсами у крыс, выращенных в сложной среде. Нейробиология обучения и памяти 65 , 48–56, https://doi.org/10.1006/nlme.1996.0005 (1996).
КАС Статья пабмед Google ученый
Исии К. и др. . In vivo динамика объема дендритных шипов в неокортексе мышей дикого типа и мышей Fmr1 KO. eNeuro 5 (2018).
Амитай, Ю. Таламокортикальные синаптические связи: эффективность, модуляция, ингибирование и пластичность. Обзоры по нейронаукам 12 , 159–174 (2001).
MathSciNet КАС Статья Google ученый
Хигашимори Х. и др. . Селективная делеция астроглиального FMRP нарушает регуляцию транспортера глутамата GLT1 и вносит вклад в фенотипы синдрома ломкой Х-хромосомы In Vivo . Journal of Neuroscience 36 , 7079–7094, https://doi.org/10.1523/JNEUROSCI.1069-16.2016 (2016).
КАС Статья пабмед Google ученый
Antoine, M.W., Langberg, T., Schnepel, P. & Feldman, D.E. Повышенное отношение возбуждения-торможения стабилизирует возбудимость синапсов и контуров в четырех моделях мышей с аутизмом. Нейрон 101 , 648–661 (2019).
КАС Статья Google ученый
Рубинштейн, Дж. Л. Р. и Мерцених, М. М. Модель аутизма: повышенное соотношение возбуждения/торможения в ключевых нервных системах. Гены, мозг и поведение 2 , 255–267 (2003).
КАС Статья Google ученый
О’Доннелл, К.
, Гонсалвес, Дж. Т., Портера-Кайо, К. и Сейновски, Т. Дж. Помимо дисбаланса возбуждения/торможения в многомерных моделях изменений нейронных цепей при заболеваниях головного мозга. Элиф 6 , e26724 (2017).Артикул Google ученый
Турко П., Гроберман К., Брова Ф., Кобб С. и Вида И. Дифференциальная зависимость ГАМКергических и глутаматергических нейронов от глии для установления синаптической передачи. Кора головного мозга (2018).
Ришер, В. К. и др. . Астроциты улучшают связность коры в дендритных шипиках. Элиф 3 , e04047 (2014).
Артикул Google ученый
 Развивающая медицина и детская неврология 44 , 724–728, https://doi.org/10.1111/j.1469-8749.2002.tb00277.x (2002).
Развивающая медицина и детская неврология 44 , 724–728, https://doi.org/10.1111/j.1469-8749.2002.tb00277.x (2002). «>
«>Джин, П., Алиш, Р. С. и Уоррен, С. Т. РНК и микроРНК при ломкой умственной отсталости Х. Nature Cell Biology 6 , 1048–1053, https://doi.org/10.1038/ncb1104-1048 (2004).
КАС Статья пабмед Google ученый
 «>
«>Контрактор А., Клячко В. А. и Портера-Кайо С. Изменение возбудимости нейронов и цепей при синдроме ломкой Х-хромосомы. Нейрон 87 , 699–715, https://doi.org/10.1016/j.neuron.2015.06.017 (2015).
КАС Статья пабмед ПабМед Центральный Google ученый
 1002/1096-8628(20010115)98:2<161::AID-AJMG1025>3.0.CO;2-B (2001).
1002/1096-8628(20010115)98:2<161::AID-AJMG1025>3.0.CO;2-B (2001). Американский журнал медицинской генетики 135 A , 155–160, https://doi.org/10.1002/ajmg.a.30709 (2005).
Американский журнал медицинской генетики 135 A , 155–160, https://doi.org/10.1002/ajmg.a.30709 (2005). «>
«>Баньи, К. и Зукин, Р. С. Синаптическая перспектива синдрома ломкой Х-хромосомы и расстройств аутистического спектра. Нейрон 101 , 1070–1088, https://doi.org/10.1016/j.neuron.2019.02.041 (2019).
КАС Статья пабмед Google ученый
 «>
«>Дейли, М.Е. и Смит, С.Дж. Динамика дендритной структуры в развивающихся срезах гиппокампа. Journal of Neuroscience 16 , 2983–94, 01.05.1996 00:01 (1996).
 Т. и др. . Синаптическая регуляция синтеза белка и хрупкого белка X. Proceedings of the National Academy of Sciences 98 , 7101–7106, https://doi.org/10.1073/pnas.141145998 (2001).
Т. и др. . Синаптическая регуляция синтеза белка и хрупкого белка X. Proceedings of the National Academy of Sciences 98 , 7101–7106, https://doi.org/10.1073/pnas.141145998 (2001). «>
«>Альварес, В. А. и Сабатини, Б. Л. Анатомическая и физиологическая пластичность дендритных шипов. Annual Review of Neuroscience 30 , 79–97, https://doi.org/10.1146/annurev.neuro.30.051606.094222 (2007).
КАС Статья пабмед Google ученый
Фремо, Р. Т. и др. . Экспрессия везикулярных переносчиков глутамата определяет два класса возбуждающих синапсов. Нейрон 31 , 247–260, https://doi.org/10.1016/S0896-6273(01)00344-0 (2001).
КАС Статья пабмед Google ученый
Wang, G. X., Smith, S. J. & Mourrain, P. Fmr1 KO и лечение фенобамом по-разному влияют на отдельные популяции синапсов неокортекса мышей. Нейрон 84 , 1273–1286, https://doi.org/10.1016/j.neuron.2014.11.016 (2014).
КАС Статья пабмед ПабМед Центральный Google ученый
Пейси, Л. К. и Доеринг, Л. С. Экспрессия FMRP в развитии в линии астроцитов: последствия для синдрома ломкой Х-хромосомы. Glia 55 , 1601–1609, https://doi.org/10.1002/glia.20573, arXiv:1505.00300v1 (2007).
Артикул Google ученый
Пэн Ф., Олдридж Г. М., Гриноу В. Т. и Ган В.-Б. Дендритная нестабильность позвоночника и нечувствительность к модуляции сенсорным опытом в мышиной модели синдрома ломкой Х-хромосомы. Proceedings of the National Academy of Sciences 107 , 17768–17773, https://doi.org/10.1073/pnas.1012496107 (2010).
ОБЪЯВЛЕНИЕ Статья Google ученый
Knott, G.W., Quairiaux, C., Genoud, C. & Welker, E. Формирование дендритных шипиков с ГАМКергическими синапсами, индуцированное стимуляцией усов у взрослых мышей. Нейрон 34 , 265–273, https://doi.org/10.1016/S0896-6273(02)00663-3 (2002).
КАС Статья пабмед Google ученый
White, E.L. Идентифицированные нейроны в коре головного мозга мыши, которые являются постсинаптическими к таламокортикальным окончаниям аксонов: комбинированное исследование Гольджи-электронной микроскопии и дегенерации. Journal of Comparative Neurology 181 , 627–661, https://doi.org/10.1002/cne.
0310 (1978).КАС Статья пабмед Google ученый
Хеллер, Дж. П., Михалюк, П., Сугао, К. и Русаков, Д. А. Исследование наноорганизации астроглии с помощью многоцветной микроскопии сверхвысокого разрешения. Journal of Neuroscience Research 95 , 2159–2171, https://doi.org/10.1002/jnr.24026 (2017).
КАС Статья пабмед Google ученый
Ючевски, К. и др. . Расширение соматосенсорной карты и измененная обработка тактильных сигналов в мышиной модели синдрома ломкой Х-хромосомы. Нейробиология болезней 96 , 201–215 (2016).
Артикул Google ученый
Скачать ссылки
Благодарности
Эта работа была поддержана грантами Национального института здравоохранения: NIH-TRA 1R01NS0, R01 Mh209475, R01 Mh204227, R01 Mh211769 NS9 и R0411. Его также поддержали: Национальный научный фонд (NSF), Управление военно-морских исследований США (ONR), Исследовательское управление армии США (ARO) и Национальное агентство геопространственной разведки (NGA). Подарки GS от Google, Microsoft и Amazon также приветствуются. Авторы благодарят профессора Ричарда Вайнберга из Университета Северной Каролины за его всегда острые идеи о синапсах.
Информация об авторе
Авторы и организации
Факультет электротехники, Университет Дьюка, Дарем, США
Аниш К. Симхал и Гильермо Сапиро
Кафедра молекулярной биологии Калифорнийского университета развития и биаллелогии Санта-Круз, США
Yi Zuo
Кафедра молекулярной и клеточной физиологии, Медицинский факультет Стэнфордского университета, Стэнфорд, США
Marc M. Perez, Daniel V. Madison и Kristina D. Micheva
Факультеты биомедицинской инженерии, компьютерных наук и математики Университета Дьюка, Дарем, США
Гильермо Сапиро
Авторы
- Аниш К. Симхал в
PubMed Google Scholar
- Yi Zuo
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Marc M. Perez
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Дэниел В. Мэдисон
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Guillermo Sapiro
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Кристина Дмитриевна Мичева
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Contributions
A.K.S. участвовал в разработке и анализе исследования, создал программное обеспечение, участвовал в написании рукописи. Ю.З. участвовал в разработке концепции исследования, интерпретации данных и написании рукописи. М.М.П. получили данные и участвовали в анализе. Д.В.М. участвовал в разработке концепции исследования. Г. С. участвовал в разработке концепции и дизайна работы, а также в написании рукописи. К.Д.М. участвовал в концепции, дизайне, анализе, интерпретации и написании.
Автор, ответственный за переписку
Аниш К. Симхал.
Декларации этики
Конкурирующие интересы
K.D.M. имеет долю учредителя в компании Aratome, LLC (Менло-Парк, Калифорния), которая производит материалы и услуги для матричной томографии. К.Д.М. также указан как изобретатель двух патентов США на методы матричной томографии, выданных Стэнфордскому университету.
Дополнительная информация
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительная информация
Дополнительные рисунки и таблицы
Права и разрешения
Открытый доступ или формате, при условии, что вы укажете автора(ов) оригинала и источник, предоставите ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатка и разрешения
Об этой статье
Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Многогранное влияние валентности антигена на состав В-клеточного ответа и дифференцировку In Vivo
. 2020 15 сентября; 53(3):548-563.e8.
doi: 10. 1016/j.immuni.2020.08.001.
Epub 2020 27 августа.
Ю Като 1 , Роберт К. Эбботт 1 , Брайан Л. Фриман 2 , Соня Хаупт 3 , Беттина Грошел 4 , Мурильо Сильва 5 , Сергей Менис 4 , Даррелл Дж. Ирвин 6 , Уильям Р. Шиф 7 , Шейн Кротти 8
Принадлежности
- 1 Центр исследований инфекционных заболеваний и вакцин, Институт иммунологии Ла-Хойя (LJI), Ла-Хойя, Калифорния , США; Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США.
- 2 Центр исследований инфекционных заболеваний и вакцин, Институт иммунологии Ла-Хойя (LJI), Ла-Хойя, Калифорния , США.
- 3 Центр исследований инфекционных заболеваний и вакцин, Институт иммунологии Ла-Хойи (LJI), Ла-Хойя, Калифорния , США; Медицинский факультет, Отделение инфекционных заболеваний и глобального общественного здравоохранения, Калифорнийский университет, Сан-Диего (UCSD), Ла-Хойя, Калифорния , США.
- 4 Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США; Департамент иммунологии и микробиологии, Научно-исследовательский институт Скриппса, Ла-Хойя, Калифорния , США; Центр нейтрализующих антител IAVI, Исследовательский институт Скриппса, Ла-Хойя, Калифорния , США.
- 5 Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США; Институт комплексных исследований рака им. Коха, Массачусетский технологический институт.
- 6 Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США; Институт комплексных исследований рака им. Коха, Массачусетский технологический институт; Департамент биологической инженерии, Массачусетский технологический институт, Кембридж, Массачусетс 02139, США; Департамент материаловедения и инженерии, Массачусетский технологический институт, Кембридж, Массачусетс 02139, США; Больница общего профиля Массачусетского института Рагона, Массачусетский технологический институт и Гарвардский университет, Кембридж, Массачусетс 02139, США; Медицинский институт Говарда Хьюза, Чеви-Чейз, Мэриленд, 20815, США.
- 7 Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США; Департамент иммунологии и микробиологии, Научно-исследовательский институт Скриппса, Ла-Хойя, Калифорния , США; Центр нейтрализующих антител IAVI, Исследовательский институт Скриппса, Ла-Хойя, Калифорния , США; Больница общего профиля Массачусетского института Рагона, Массачусетский технологический институт и Гарвардский университет, Кембридж, Массачусетс 02139, США. Электронный адрес: [email protected].
- 8 Центр исследований инфекционных заболеваний и вакцин, Институт иммунологии Ла-Хойи (LJI), Ла-Хойя, Калифорния , США; Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США; Медицинский факультет, Отделение инфекционных заболеваний и глобального общественного здравоохранения, Калифорнийский университет, Сан-Диего (UCSD), Ла-Хойя, Калифорния , США. Электронный адрес: [email protected].
- PMID: 32857950
- PMCID: PMC7451196
- DOI: 10.1016/j.иммуни.2020.08.001
Бесплатная статья ЧВК
Ю Като и соавт. Иммунитет. .
Бесплатная статья ЧВК
. 2020 15 сентября; 53(3):548-563.e8.
doi: 10.1016/j.immuni.2020.08.001.
Epub 2020 27 августа.
Авторы
Ю Като 1 , Роберт К. Эбботт 1 , Брайан Л. Фриман 2 , Соня Хаупт 3 , Беттина Грошел 4 , Мурильо Сильва 5 , Сергей Менис 4 , Даррелл Дж. Ирвин 6 , Уильям Р. Шиф 7 , Шейн Кротти 8
Принадлежности
- 1 Центр исследований инфекционных заболеваний и вакцин, Институт иммунологии Ла-Хойя (LJI), Ла-Хойя, Калифорния , США; Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США.
- 2 Центр исследований инфекционных заболеваний и вакцин, Институт иммунологии Ла-Хойя (LJI), Ла-Хойя, Калифорния , США.
- 3 Центр исследований инфекционных заболеваний и вакцин, Институт иммунологии Ла-Хойи (LJI), Ла-Хойя, Калифорния г., США; Медицинский факультет, Отделение инфекционных заболеваний и глобального общественного здравоохранения, Калифорнийский университет, Сан-Диего (UCSD), Ла-Хойя, Калифорния , США.
- 4 Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США; Департамент иммунологии и микробиологии, Научно-исследовательский институт Скриппса, Ла-Хойя, Калифорния , США; Центр нейтрализующих антител IAVI, Исследовательский институт Скриппса, Ла-Хойя, Калифорния , США.
- 5 Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США; Институт комплексных исследований рака им. Коха, Массачусетский технологический институт.
- 6 Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США; Институт комплексных исследований рака им. Коха, Массачусетский технологический институт; Департамент биологической инженерии, Массачусетский технологический институт, Кембридж, Массачусетс 02139, США; Департамент материаловедения и инженерии, Массачусетский технологический институт, Кембридж, Массачусетс 02139, США; Больница общего профиля Массачусетского института Рагона, Массачусетский технологический институт и Гарвардский университет, Кембридж, Массачусетс 02139, США; Медицинский институт Говарда Хьюза, Чеви-Чейз, Мэриленд, 20815, США.
- 7 Консорциум Скриппса по разработке вакцин против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния , США; Отделение иммунологии и микробиологии, Научно-исследовательский институт Скриппса, Ла-Хойя, Калифорния г., США; Центр нейтрализующих антител IAVI, Исследовательский институт Скриппса, Ла-Хойя, Калифорния , США; Больница общего профиля Массачусетского института Рагона, Массачусетский технологический институт и Гарвардский университет, Кембридж, Массачусетс 02139, США. Электронный адрес: [email protected].
- 8 Центр исследований инфекционных заболеваний и вакцин, Институт иммунологии Ла-Хойи (LJI), Ла-Хойя, Калифорния , США; Консорциум Скриппса по разработке вакцины против ВИЧ/СПИДа (CHAVD), Ла-Хойя, Калифорния г., США; Медицинский факультет, Отделение инфекционных заболеваний и глобального общественного здравоохранения, Калифорнийский университет, Сан-Диего (UCSD), Ла-Хойя, Калифорния , США. Электронный адрес: [email protected].
- PMID: 32857950
- PMCID: PMC7451196
- DOI: 10.1016/j.иммуни.2020.08.001
Абстрактный
Как валентность антигена влияет на В-клетки in vivo во время иммунного ответа, не совсем понятно. Здесь, используя иммуногены ВИЧ с определенной валентностью в диапазоне от 1 до 60, мы исследовали роль валентности антигена во время различных фаз ответа В-клеток in vivo. Иммуногены с высокой степенью мультимеризации предпочтительно быстро активируют родственные В-клетки с небольшой дискриминацией по аффинности. Это привело к сильной ранней индукции факторов транскрипции IRF4 (фактор регуляции интерферона 4) и Bcl6, управляя как ранними экстрафолликулярными реакциями плазматических клеток, так и реакциями зародышевых центров в CD4 + Т-клеточно-зависимый, с участием В-клеток с широким диапазоном сродства. Антигены с низкой валентностью индуцировали меньшие ответы эффекторных В-клеток с преимущественным привлечением высокоаффинных В-клеток. Таким образом, валентность антигена оказывает многогранное влияние на ответ В-клеток и может определять пороги аффинности и конкурентные ландшафты для В-клеток in vivo, что имеет значение для разработки вакцины.
Ключевые слова: В-клетка; ВИЧ; ВРК01; порог близости; зародышевый центр; нацеливание на зародышевую линию; наночастицы; плазматическая клетка; вакцина; валентность.
Copyright © 2020 Elsevier Inc. Все права защищены.
Заявление о конфликте интересов
Декларация об интересах С.М. и W.R.S. являются изобретателями патентных заявок, поданных IAVI и Scripps на eOD-GT8 60-mer.
Цифры
Графический реферат
Графический реферат
Рисунок 1
Валентность антигена влияет на родственный B…
Рисунок 1
Антигенные валентности влияют на стимуляцию родственных В-клеток In Vivo в течение нескольких часов (A) В масштабе…
Фигура 1 Валентность антигена влияет на родственную стимуляцию В-клеток In Vivo в течение нескольких часов (A) Масштабные модели конструкций eOD-GT5: eOD (зеленый), CD4bs (желтый), каркас (голубой) и гликаны (сланцевый). (B–D) Прижизненная визуализация iLN через 2–3,5 часа после иммунизации антигенами eOD-GT5. (Б) Схема. (C) Скорость В-клеток VRC01 gHL , нормированная на поликлональные В-клетки. (D) Стационарные В-клетки VRC01 gHL (скорость <3 мкм/мин). Данные в (A)–(D) взяты из трех экспериментов с n = 2–5 мышей в группе. Бары представляют среднее значение. (Э–Н) VRC01 gHL Активация В-клеток через 24 часа. (E) Маркеры активации на В-клетках VRC01 gHL . (F) Количественная оценка маркеров активации в (E), средняя геометрическая интенсивность флуоресценции (MFI). (G) Размер клетки VRC01 gHL B. (H) Количественная оценка размера клеток VRC01 gHL B (среднее значение FSC-A). Данные в (E)–(H) взяты из двух экспериментов с n = 6 мышами. Бары представляют среднее значение. ∗ р < 0,05; ∗∗ р < 0,01; ∗∗∗ р < 0,001; ∗∗∗∗ р < 0,0001. См. также рисунки S1 и S2, видео S1 и таблицу S1.
Рисунок 2
Дифференциальные эффекты антигенных валентностей…
Рисунок 2
Дифференциальные эффекты валентности антигена на деление В-клеток In Vivo (A–C) Пролиферативный…
Фигура 2 Дифференциальные эффекты валентностей антигена на деление В-клеток In Vivo (A–C) Пролиферативный ответ В-клеток VRC01 gHL через 3 дня после иммунизации. (A) Профили дивизий. Ворота показывают клетки CTV hi (неразделенные) и CTV lo (разделенные). (B) Разделенный номер клетки VRC01 gHL B. Столбцы представляют среднее геометрическое. (C) Неделящиеся В-клетки VRC01 gHL как частота общих В-клеток, нормализованная по отношению к неиммунизированному контролю. Бары представляют среднее значение. Данные в (A)–(C) взяты из двух экспериментов с n = 6 мышами. (D–F) VRC01 gHL Характер активации и локализации В-клеток через 24 ч после иммунизации. (D) Крупные (FSC-A hi ) CCR7 hi VRC01 gHL В-клетки. (E) Репрезентативные iLN, показывающие В-клетки VRC01 gHL (CTV, белый), зоны В-клеток (IgD, синий) и зоны Т-клеток (CD4, зеленый). Сигналы CTV были расширены цифровым способом. Масштабная линейка, 300 мкм. (F) Распределение В-клеток VRC01 gHL . Общее количество проанализированных клеток указано в столбцах. Данные в (D)–(F) взяты из двух экспериментов с n = 6 мышами. ∗ р < 0,05; ∗∗ р < 0,01; ∗∗∗ р < 0,001; ∗∗∗∗ р < 0,0001.
Рисунок 3
TD Индукция ранней ГХ…
Рисунок 3
TD Индукция ранних GC B-клеток и PC (A)–(F) Эффекты CD4…
Рисунок 3 TD Индукция ранних GC B-клеток и PC (A)–(F) Влияние истощения CD4 + Т-клеток на VRC01 gHL В-клеточные ответы на 60-член eOD-GT5. (A) Профили дивизий. На воротах показаны неделимые и разделенные клетки. (B) Разделенное число клеток VRC01 gHL B, нормализованное по контрольной группе с 60 мерами. (C) Неделящиеся В-клетки VRC01 gHL как частота общих В-клеток, нормализованная по отношению к неиммунизированному контролю. Данные в (A)–(C) взяты из четырех экспериментов с n = 6–11 мышей. Бары представляют среднее значение. (D) Экспрессия Bcl6 и IRF4 с помощью разделенного VRC01 gHL (красный) и эндогенные (серый) В-клетки. Числа в квадрантах представляют собой проценты. (E) Разделенные В-клетки VRC01 gHL , которые представляют собой В-клетки Bcl6 hi IRF4 lo GC. (F) Разделенные В-клетки VRC01 gHL , которые представляют собой IRF4 hi Bcl6 lo шт. Данные в (D)–(F) взяты из двух экспериментов с n = 5–6 мышами. Бары представляют среднее значение. (G)–(L) Пролиферация и дифференцировка В-клеток VRC01 gHL , индуцированная 4-мерным или 60-мерным eOD-GT5 gp61 на 4-й день. (G) Профили деления. На воротах показаны неделимые и разделенные клетки. (H) Разделенный VRC01 gHL Номер ячейки B. Столбцы представляют среднее геометрическое. (I) Неделящиеся В-клетки VRC01 gHL как частота общих В-клеток, нормализованная по отношению к неиммунизированному контролю. Бары представляют среднее значение. (J) Экспрессия Bcl6 и IRF4 в разделенных В-клетках VRC01 gHL после иммунизации (синий или красный) и неразделенных В-клетках VRC01 gHL от наивных мышей (серый). Проценты указаны в квадрантах. (K) Разделившиеся VRC01 gHL В-клетки, представляющие собой Bcl6 hi IRF4 lo GC В-клетки. (L) Разделенные В-клетки VRC01 gHL , которые представляют собой IRF4 hi Bcl6 lo шт. Данные в (G)–(L) взяты из двух экспериментов с n = 3–5 мышей. (М) Разделенные В-клетки VRC01 gHL , которые представляют собой В-клетки Bcl6 hi раннего GC (фиолетовые), IRF4 hi PCs (зеленые), клетки Bc6 hi IRF4 hi (черные) или двойные отрицательные ( белый). «d0» представляет собой неразделенные В-клетки VRC01 gHL от интактных мышей. Данные взяты из двух-четырех экспериментов с n = 2–8 мышей. ∗∗ р < 0,01; ∗∗∗∗ р < 0,0001. См. также рисунок S3.
Рисунок 4
Высокомультимеризованный низкоаффинный антиген Эффективно…
Рисунок 4
Высокомультимеризованный низкоаффинный антиген эффективно индуцирует ранние В-клеточные ответы (A и B)…
Рисунок 4 Высокомультимеризованный низкоаффинный антиген эффективно индуцирует ранние В-клеточные ответы (A и B) Прижизненная визуализация iLN через 2–3,5 часа после иммунизации 60-мерным eOD-GT2. (A) Нормализованная скорость клеток VRC01 gHL B ( ∗∗∗∗ p <0,0001 по сравнению с неиммунизированными). (B) Частота стационарных В-клеток VRC01 gHL (скорость <3 мкм/мин; * p <0,05 по сравнению с неиммунизированными). Данные в (A) и (B) взяты из трех экспериментов с n = 4 мышами. Бары представляют среднее значение. См. также рисунки 1C и 1D. (С и D) VRC01 gHL Активация В-клеток через 24 часа. (C) Размер клетки VRC01 gHL B (средний FSC-A). (D) Маркеры активации на В-клетках VRC01 gHL (нормализованный геометрический MFI). Данные в (C) и (D) взяты из трех экспериментов с n = 6–9 мышей. Бары представляют среднее значение. (E) Распределение В-клеток VRC01 gHL через 24 часа. Данные взяты из двух экспериментов с n = 6 мышами и 580 клетками. (F)–(H) Прижизненная визуализация, показывающая взаимодействие Т- и В-клеток через 24 часа. (F) Схема. (G) Репрезентативные кадры фильма. (Н) VRC01 В-клетки gHL (ряды) были классифицированы как «в контакте» (красный) или «не в контакте» (синий) с Т-клетками SMARTA CD4 + . Показаны случайно выбранные 71 VRC01 gHL В-клетки. (I) Процент времени, в течение которого В-клетки VRC01 gHL находились в контакте. Данные в (G) – (I) взяты из двух экспериментов с n = 3 мышами. (J – P) Пролиферация и дифференцировка В-клеток VRC01 gHL через 3 дня после иммунизации. (J) Профили дивизий. Ворота идентифицируют неразделенные и разделенные клетки. (K) CTV (геометрический MFI) на разделенном VRC01 gHL В-клетки, нормированные к eOD-GT5 gp61 60-мер. (L) Разделенный номер клетки VRC01 gHL B. (M) Неделящиеся В-клетки VRC01 gHL как частота общих В-клеток, нормализованных к неиммунизированным. (N) Экспрессия Bcl6 и IRF4 в разделенных В-клетках VRC01 gHL . Проценты указаны в квадрантах. (O) Разделенные В-клетки VRC01 gHL , которые представляют собой В-клетки Bcl6 hi IRF4 lo GC. (P) Разделившиеся VRC01 gHL В-клетки, которые представляют собой IRF4 hi Bcl6 lo шт. Данные в (J)–(P) взяты из двух экспериментов с n = 6 мышами. Бары представляют среднее значение. ∗ р < 0,05; ∗∗ р < 0,01; ∗∗∗ р < 0,001; ∗∗∗∗ р < 0,0001. См. также видео S2 и S3 и рисунок S4.
Рисунок 5
Валентность антигена влияет на величину…
Рисунок 5
Валентность антигена влияет на величину и состав ответа GC В-клеток…
Рисунок 5 Валентность антигена влияет на величину и состав ответов GC B-клеток (A–L) VRC01 gHL и эндогенных ответов GC B-клеток через 6 дней после иммунизации. (А) Схема. (B) Общий ответ GC B-клеток (слева) и ответ VRC01 gHL GC B-клеток (справа). Указаны проценты. (C) Общее количество клеток GC B. Полосы указывают среднее значение. (Г) VRC01 gHL В-клетки GC в процентах от общего количества В-клеток GC. Полосы указывают среднее геометрическое. (E) Номер ячейки VRC01 gHL GC B. Полосы указывают среднее значение. Данные в (A)–(E) взяты из трех экспериментов с n = 9–10 мышей в группе. (F) Репрезентативные iLN: В-клетки VRC01 gHL (GFP, зеленый), IgD (синий), GL7 (красный) и Т-клетки SMARTA CD4 + (CD45.1, белый). GC представляют собой область GL7 + IgD — . Желтые квадраты показывают примеры GC. Красный квадрат показывает экстрафолликулярный VRC01 gHL В-клетки. Масштабные линейки, 300 мкм. (G) площадь GC на секцию. Полосы указывают среднее значение. (H) VRC01 gHL Плотность клеток B в GC (количество на мм 2 ). Линии в первом квартиле, медиане и третьем квартиле; n = 36 GC (4-мер) и 70 GC (60-мер). (I) VRC01 gHL GC Количество клеток В на секцию. Полосы указывают среднее значение. Данные в (F)–(I) взяты из двух экспериментов с n = 6 мышами. (J) Эндогенные В-клетки GC. Ворота идентифицируют «высокоаффинные» клетки с обнаруживаемой аффинностью к антигенным зондам. Указаны проценты. (K) Частота высокоаффинных эндогенных В-клеток GC. (L) Количество высокоаффинных эндогенных В-клеток GC. Данные в (J)–(L) взяты из двух экспериментов с n = 7 мышами. Полосы указывают среднее значение. (М–О) VRC01 gHL GC В-клеточные ответы через 6 дней после иммунизации eOD-GT2 gp61 4-мерный или 60-мерный. (M) Общее количество клеток GC B. Линиями показаны данные для eOD-GT5 gp61 4-мер (синий) и 60-мер (красный). См. также рисунок 5C. (N) В-клетки VRC01 gHL в процентах от общего количества В-клеток GC. Линиями показаны данные для eOD-GT5 gp61 4-мер (синий) и 60-мер (красный). См. также рисунок 5D. (O) VRC01 gHL GC Номер ячейки B. Линиями показаны данные для eOD-GT5 gp61 4-мер (синий) и 60-мер (красный). См. также рисунок 5E. Данные в (M) – (O) взяты из двух экспериментов с n = 7 мышами. Столбцы указывают среднее значение (M) или среднее геометрическое (N и O). ∗ р < 0,05; ∗∗ р < 0,01; ∗∗∗ р < 0,001; ∗∗∗∗ р < 0,0001. См. также рисунок S5.
Рисунок 6
Валентность антигена влияет на величину…
Рисунок 6
Валентность антигена влияет на величину и состав экстрафолликулярных ответов ПК (A–I)…
Рисунок 6 Валентность антигена влияет на величину и состав внефолликулярных ответов ПК (A–I) VRC01 gHL и эндогенных ответов ПК через 6 дней после иммунизации eOD-GT5 gp61 4-мерный или 60-мерный. (A) Всего ответов ПК (слева) и VRC01 gHL Ответы ПК (справа). (B) Общее количество ПК. Полосы указывают среднее значение. (C) Клетки VRC01 gHL в процентах от общего числа ПК. Столбцы указывают медиану. (D) VRC01 gHL Номер ПК. Столбцы указывают медиану. Данные в (A)–(D) взяты из трех экспериментов с n = 9–10 мышами. (E) CD4bs-специфические ответы IgG (среднее геометрическое ± геометрическая стандартная ошибка). 2 эксперимента, n = 7. (F) Эндогенные ПК (IgD – CD138 + ). Ворота идентифицируют «высокоаффинные» клетки с обнаруживаемой аффинностью к антигенным зондам. (G) Высокоаффинные эндогенные ПК. (H и I) Высокоаффинные эндогенные GC B-клетки и PC после иммунизации (H) eOD-GT5 gp61 4-мер или (I) 60-мер. См. Рисунок 5K. (J – L) VRC01 gHL PC ответы через 6 дней после иммунизации eOD-GT2 gp61 4-мер или 60-мер. (J) Общее количество ПК. Полосы указывают среднее значение. Линиями показаны данные для eOD-GT5 gp61 4-мер (синий) и 60-мер (красный). (K) Клетки VRC01 gHL в процентах от общего числа ПК. Столбцы указывают медиану. Линиями показаны данные для eOD-GT5 gp61 4-мер (синий) и 60-мер (красный). (L) VRC01 gHL Номер ПК. Столбцы указывают медиану. Линиями показаны данные для eOD-GT5 gp61 4-мер (синий) или 60-мер (красный). Данные в (F)–(L) взяты из двух экспериментов (n = 6). ∗ р < 0,05; ∗∗ р < 0,01; ∗∗∗ р < 0,0001; ∗∗∗∗ р < 0,0001. См. также рисунок S5.
Рисунок 7
Постоянство ячеек VRC01-класса B…
Рисунок 7
Персистенция клеток VRC01-класса B в GCs (A–F) Кинетика VRC01 gHL GC…
Рисунок 7 Персистенция клеток VRC01-класса B в GCs (A–F) Кинетика VRC01 gHL GC B-клеточные ответы после иммунизации антигенами eOD-GT5 gp61 (A–D) или eOD-GT2 gp61 (A и Э-Г). (A) Общее количество клеток GC B. Объединенные данные антигенов eOD-GT2 gp61 и eOD-GT5 gp61 . (B и E) Клетки VRC01 gHL в процентах от общего количества В-клеток GC (среднее геометрическое ± геометрическая стандартная ошибка). (С и F) VRC01 gHL GC Номер В-клетки (среднее геометрическое ± геометрическая стандартная ошибка). (D и G) Кумулятивный ответ В-клеток VRC01 gHL GC. (D) Площадь под кривой (AUC) логарифмически преобразованных данных в (C). (G) AUC логарифмически преобразованных данных в (F). Среднее + стандартное отклонение. Данные в (A)–(G) взяты из двух экспериментов, n = 7, ∗ p <0,05; ∗∗ р < 0,01; ∗∗∗ p < 0,001, ∗∗∗∗ p < 0,0001. См. также рисунок S6.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
BRAD4 играет критическую роль в ответе зародышевого центра, регулируя активацию Bcl-6 и NF-kB.
Гао Ф, Ян И, Ван З, Гао С, Чжэн Б. Гао Ф. и др. Клеточный Иммунол. 2015 март; 294(1):1–8. doi: 10.1016/j.cellimm.2015.
01.010. Epub 2015 28 января.
Клеточный Иммунол. 2015.
PMID: 25656449Многомасштабное моделирование зародышевого центра резюмирует временной переход от В-клеток памяти к дифференцировке плазматических клеток, регулируемый с помощью клеток Tfh на основе аффинности антигена.
Мерино Техеро Э., Лашгари Д., Гарсия-Валиенте Р., Гао X, Краусте Ф., Роберт П.А., Мейер-Херманн М., Мартинес М.Р., ван Хэм С.М., Гикема Дж.Е., Хофслут Х., ван Кампен АХК. Меринос Техеро Э. и др. Фронт Иммунол. 2021 5 фев; 11:620716. doi: 10.3389/fimmu.2020.620716. Электронная коллекция 2020. Фронт Иммунол. 2021. PMID: 33613551 Бесплатная статья ЧВК.
Направленная Thpok транскрипционная схема способствует экспрессии Bcl6 и Maf для организации дифференцировки Т-фолликулярных хелперов.
Vacchio MS, Ciucci T, Gao Y, Watanabe M, Balmaceno-Criss M, McGinty MT, Huang A, Xiao Q, McConkey C, Zhao Y, Shetty J, Tran B, Pepper M, Vahedi G, Jenkins MK, McGavern ДБ, Босселут Р. Ваккио М.С. и др. Иммунитет. 2019 17 сентября; 51(3):465-478.e6. doi: 10.1016/j.immuni.2019.06.023. Epub 2019 15 августа. Иммунитет. 2019. PMID: 31422869 Бесплатная статья ЧВК.
Фактор транскрипции IRF4 определяет формирование зародышевого центра посредством дифференцировки фолликулярных Т-хелперов.
Боллиг Н., Брюстле А., Келлнер К., Акерманн В., Абасс Э., Райфер Х., Камара Б., Брендель С., Гил Г., Ботур Э., Хубер М., Пол С., Элли А., Крочек Р.А., Нуриева Р., Донг С., Джейкоб Р., Мак Т.В., Лохофф М. Боллиг Н. и др. Proc Natl Acad Sci U S A. 2012 г., 29 мая.;109(22):8664-9. doi: 10.1073/pnas.1205834109. Epub 2012 2 мая. Proc Natl Acad Sci U S A.
2012.
PMID: 22552227
Бесплатная статья ЧВК.Регуляторный фактор интерферона 4 и 8 в развитии В-клеток.
Лу Р. Лу Р. Тренды Иммунол. 2008 окт; 29 (10): 487-92. doi: 10.1016/jit.2008.07.006. Epub 2008 3 сентября. Тренды Иммунол. 2008. PMID: 18775669 Бесплатная статья ЧВК. Обзор.
Посмотреть все похожие статьи
Цитируется
Усиление гуморального ответа за счет мультивалентного антигенного дисплея на тимус-независимых ДНК-оригами каркасах.
Вамхофф Э.К., Ронсард Л., Фельдман Дж., Хаузер Б.М., Кнаппе Г.А., Романов А., Лам Э., Сен-Дени К., Балаш А.Б., Шмидт А., Лингвуд Д., Бате М. Wamhoff EC и соавт. bioRxiv. 2022, 24 августа: 2022.08.16.
504128. дои: 10.1101/2022.08.16.504128. Препринт.
bioRxiv. 2022.
PMID: 36032975
Бесплатная статья ЧВК.Антитела от первичных гуморальных ответов модулируют рекрутирование наивных В-клеток во время вторичных ответов.
Tas JMJ, Koo JH, Lin YC, Xie Z, Steichen JM, Jackson AM, Hauser BM, Wang X, Cottrell CA, Torres JL, Warner JE, Kirsch KH, Weldon SR, Groschel B, Nogal B, Ozorowski G, Бангару С., Фелпс Н., Адачи Ю., Эскандарзаде С., Кубиц М., Бертон Д.Р., Лингвуд Д., Шмидт А.Г., Наир У., Уорд А.Б., Шиф В.Р., Батиста Ф.Д. Тас Дж.М.Дж. и др. Иммунитет. 4 августа 2022 г .: S1074-7613 (22) 00355-7. doi: 10.1016/j.immuni.2022.07.020. Онлайн перед печатью. Иммунитет. 2022. PMID: 35987201 Бесплатная статья ЧВК.
IL-21 играет критическую роль в создании зародышевых центров путем усиления ранней пролиферации В-клеток.
Дворшек А.Р., Маккензи С.И., Робинсон М.Дж., Дин З., Питт С., О’Доннелл К., Зотос Д., Бринк Р., Тарлинтон Д.М., Кваст И. Дворчек А.Р. и соавт. Отчет EMBO, 5 сентября 2022 г .; 23 (9): e54677. doi: 10.15252/embr.202254677. Epub 2022 8 июля. Представитель EMBO 2022 г. PMID: 35801309Бесплатная статья ЧВК.
Иммунизация самособирающейся вакциной на основе наночастиц, отображающей EBV gH/gL, защищает гуманизированных мышей от летального заражения вирусом.
Малхи Х., Хомад Л.Дж., Ван Ю.Х., Пудел Б., Фиала Б., Борст А.Дж., Ван Д.Й., Уолки С., Прайс Дж., Уолл А., Сингх С., Муди З., Картер Л., Ханда С., Корренти К.Э., Стоддард Б.Л., Вислер Д., Панцера М., Олсон Дж., Кинг Н.П., Макгуайр А.Т. Малхи Х. и др. Сотовый представитель Мед. 2022 21 июня; 3 (6): 100658. doi: 10.1016/j.xcrm.2022.100658. Epub 2022 14 июня. Сотовый представитель Мед.
2022.
PMID: 35705092
Бесплатная статья ЧВК.Иммуноген наночастиц домена III вируса Powassan вызывает нейтрализующие и защитные антитела у мышей.
Малонис Р.Дж., Георгиев Г.И., Хаслвантер Д., ВанБларган Л.А., Фэллон Г., Верньолл О., Кэхилл С.М., Харрис Р., Коуберн Д., Чандран К., Даймонд М.С., Лай Дж.Р. Малонис Р.Дж. и соавт. PLoS Патог. 9 июня 2022 г .; 18 (6): e1010573. doi: 10.1371/journal.ppat.1010573. электронная коллекция 2022 июнь. PLoS Патог. 2022. PMID: 35679349 Бесплатная статья ЧВК.
Просмотреть все статьи «Цитируется по»
использованная литература
- Abbott R.K., Lee J.H., Menis S., Skog P., Rossi M., Ota T., Kulp D.W., Bhullar D., Kalyuzhniy O., Havenar-Daughton C. Частота и сродство предшественников определяют конкурентоспособность B-клеток в зародышевых центрах , протестированы с иммуногенами вакцины против ВИЧ, нацеленными на зародышевую линию. Иммунитет. 2018;48:133–146.e6.
—
ЧВК
—
пабмед
- Abbott R.K., Lee J.H., Menis S., Skog P., Rossi M., Ota T., Kulp D.W., Bhullar D., Kalyuzhniy O., Havenar-Daughton C. Частота и сродство предшественников определяют конкурентоспособность B-клеток в зародышевых центрах , протестированы с иммуногенами вакцины против ВИЧ, нацеленными на зародышевую линию.
- Аллен К.Д.К., Окада Т., Систер Дж.Г. Организация зародышевого центра и клеточная динамика. Иммунитет. 2007; 27: 190–202. — ЧВК — пабмед
- Бахманн М. Ф., Рорер У.Х., Кюндиг Т.М., Бюрки К., Хенгартнер Х., Цинкернагель Р.М. Влияние организации антигена на реактивность В-клеток. Наука. 1993; 262:1448–1451.
—
пабмед
- Бахманн М.
- Бахманн М.Ф., Цинкернагель Р.М. Влияние структуры вируса на ответы антител и формирование серотипа вируса. Иммунол. Сегодня. 1996; 17: 553–558. — пабмед
- Батиста Ф.Д., Ибер Д., Нойбергер М.С. В-клетки приобретают антиген от клеток-мишеней после образования синапса. Природа. 2001;411:489–494. — пабмед
Типы публикаций
термины MeSH
вещества
Грантовая поддержка
- UM1 AI144462/AI/NIAID NIH HHS/США
- UM1 AI100663/AI/NIAID NIH HHS/США
- S10 RR027366/RR/NCRR NIH HHS/США
- P01 AI048240/AI/NIAID NIH HHS/США
- K99 AI145762/AI/NIAID NIH HHS/США
- T32 AI125179/AI/NIAID NIH HHS/США
- S10 OD021831/OD/NIH HHS/США
Многогранные изменения синаптического состава и участие астроцитов в мышиной модели синдрома ломкой Х-хромосомы
. 2019 25 сентября; 9 (1): 13855.
doi: 10.1038/s41598-019-50240-x.
Аниш К Симхал 1 , И Цзо 2 , Марк М. Перес 3 , Дэниел В Мэдисон 3 , Гильермо Сапиро 4 5 , Кристина Д Мичева 3
Принадлежности
- 1 Факультет электротехники, Университет Дьюка, Дарем, США. [email protected].
- 2 Кафедра молекулярной, клеточной и биологии развития, Калифорнийский университет, Санта-Крус, США.
- 3 Кафедра молекулярной и клеточной физиологии, Медицинская школа Стэнфордского университета, Стэнфорд, США.
- 4 Факультет электротехники, Университет Дьюка, Дарем, США.
- 5 Факультеты биомедицинской инженерии, компьютерных наук и математики Университета Дьюка, Дарем, США.
- PMID: 31554841
- PMCID: PMC6761194
- DOI: 10.1038/с41598-019-50240-х
Бесплатная статья ЧВК
Аниш К. Симхал и др.
Научный представитель .
Бесплатная статья ЧВК
. 2019 25 сентября; 9 (1): 13855.
doi: 10.1038/s41598-019-50240-x.
Авторы
Аниш К Симхал 1 , И Цзо 2 , Марк М. Перес 3 , Дэниел В Мэдисон 3 , Гильермо Сапиро 4 5 , Кристина Д Мичева 3
Принадлежности
- 1 Факультет электротехники, Университет Дьюка, Дарем, США. [email protected].
- 2 Кафедра молекулярной, клеточной и биологии развития, Калифорнийский университет, Санта-Крус, США.
- 3 Кафедра молекулярной и клеточной физиологии, Медицинская школа Стэнфордского университета, Стэнфорд, США.
- 4 Факультет электротехники, Университет Дьюка, Дарем, США.
- 5 Факультеты биомедицинской инженерии, компьютерных наук и математики Университета Дьюка, Дарем, США.
- PMID: 31554841
- PMCID: PMC6761194
- DOI: 10.1038/с41598-019-50240-х
Абстрактный
Известно, что синдром ломкой Х-хромосомы (FXS), распространенная наследственная форма умственной отсталости, изменяет цепи неокортекса. Однако его влияние на различные типы синапсов, составляющих эти цепи, или на участие астроцитов, недостаточно известно. Мы использовали иммунофлуоресцентную матричную томографию для количественной оценки различных синаптических популяций и их связи с астроцитами в слоях с 1 по 4 соматосенсорной коры взрослых мышей модели FXS, мышей с нокаутом FMR1. Собранные многоканальные данные содержали приблизительно 1,6 миллиона синапсов, которые были проанализированы с помощью вероятностного детектора синапсов. Наше исследование выявляет сложные изменения синаптического типа и слоев в неокортикальной схеме мышей с нокаутом FMR1. Мы сообщаем об увеличении количества малых глутаматергических синапсов VGluT1 в слое 4, сопровождаемом уменьшением количества больших синапсов VGluT1 в слоях 1 и 4. Синапсы VGluT2 демонстрируют довольно последовательное снижение плотности в слоях 1 и 2/3. Во всех слоях мы наблюдаем потерю крупных тормозных синапсов. Наконец, уменьшается астроцитарная ассоциация возбуждающих синапсов. Способность анализировать дефицит цепи по типу синапса и вовлечению астроцитов будет иметь решающее значение для понимания того, как эти изменения влияют на функцию цепи, и, в конечном итоге, для определения целей для терапевтического вмешательства.
Заявление о конфликте интересов
К.Д.М. имеет долю учредителя в компании Aratome, LLC (Менло-Парк, Калифорния), которая производит материалы и услуги для матричной томографии. К.Д.М. также указан как изобретатель двух патентов США на методы матричной томографии, выданных Стэнфордскому университету.
Цифры
Рисунок 1
Схема запроса. В…
Рисунок 1
Схема запроса. На карикатуре слева изображен родственник…
Схема запроса. На рисунке слева показано относительное пространственное расположение различных флуоресцентных маркеров, используемых для обнаружения возбуждающего синапса, экспрессирующего VGluT1, рядом с отростком астроцита. Это визуальное описание синапса преобразуется в запрос, показанный справа от большой черной стрелки. Запрос — это определяемое пользователем описание того, как должен «выглядеть» интересующий тип синапса. При этом ожидается, что пресинаптические белковые маркеры — синапсин и VGluT1 будут колокализоваться (занимать одно и то же трехмерное пространство) друг с другом. Кроме того, ожидается, что пресинаптические, постсинаптические и астроцитарные маркеры (как группа) будут находиться рядом друг с другом. В верхней правой части рисунка показаны три 1,5 мкм × 1,5 мкм вырезки различных комбинаций маркеров, показывающих, как выглядит запрос в данных. На первом вырезе показаны синапсин и пунктум PSD-95, наложенные друг на друга; вторая вырезка включает пунктум GS, а третий вырез включает пунктум VGluT1.
Рисунок 2
Конвейер вероятностного обнаружения синапсов. В…
Рисунок 2
Конвейер вероятностного обнаружения синапсов. В первой колонке показан необработанный PSD-95 и синапсин…
фигура 2 Конвейер вероятностного обнаружения синапсов. В первом столбце показаны необработанные данные PSD-95 и синапсин. Во втором столбце показаны выходные данные метода обнаружения синапсов, где значение в каждом пикселе — это вероятность того, что пиксель принадлежит указанному определению синапса. В третьем столбце показаны результаты пороговой обработки выходной карты вероятностей. Из нашей прошлой работы мы определили эмпирический порог 0,9.быть оптимальным значением. В четвертом столбце показаны обнаружения (белым цветом), наложенные на суперпозицию данных PSD-95 и синапсов. Для этой визуализации определением синапса было смежностью PSD-95 и синапсина-пунктума минимального размера 0,2 мкм × 0,2 мкм × 0,07 мкм , что соответствует 2 PX × 2 PX PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX × 2 PX, что соответствует 2 PX . × 1 срез . Смежность определяется как две точки, занимающие непосредственно соседние пиксели или разделяющие небольшую долю пикселей.
Рисунок 3
Обзор наборов данных. (…
Рисунок 3
Обзор наборов данных. ( A ) Иммунофлюоресцентная матричная томография дикого типа и…
Рисунок 3 Обзор наборов данных. ( A ) Иммунофлуоресцентная матричная томография соматосенсорной коры мышей дикого типа и мышей с нокаутом FMR1. На левой панели показана область изображения в каждом образце, состоящая из четырех плиток, охватывающих слои коры с 1 по 4. Окрашивание ядер DAPI, объемная реконструкция 30 серийных срезов по 70 нм каждый. На правой панели показана иммунофлуоресценция синаптических (синапсин, VGluT1, VGluT2 и GAD2) и глиальных (GS) маркеров в слое 4 соматосенсорной коры мышей дикого типа и нокаутных мышей, объемная реконструкция 10 серийных срезов, 70 нм каждый. ( B ) Сводка изменений плотности пунктата в одном канале между мышами дикого типа и мышами с нокаутом с усреднением всех слоев. Графики показывают процентную разницу между мышами дикого типа и мышами с нокаутом и имеют цветовую кодировку, чтобы указать направление изменения и статистическую значимость. ( C ) На этом графике показано распределение плотности точек GS по размеру. «Маленькие» точки охватывают один срез, «средние» точки охватывают два среза, а «большие» точки охватывают три или более срезов.
Рисунок 4
Обзор плотности синапсов дикого типа…
Рисунок 4
Обзор распределения плотности синапсов дикого типа. ( A ) Верх, мультиплексная иммунофлуоресценция из…
Рисунок 4 Обзор распределения плотности синапсов дикого типа. ( A ) Вверху, мультиплексная иммунофлуоресценция из той же области одного среза соматосенсорной коры мыши дикого типа. Ядерное пятно DAPI окрашено в голубой цвет. Белыми квадратами выделены отдельные синапсы, которые на А внизу показаны с большим увеличением на 4 последовательных последовательных срезах. ( B ) Распределение типов синапсов по разным слоям. Слои со значительными различиями для типа синапса отмечены. Хотя нет существенной разницы в плотности слоев для глутаматергических синапсов в целом и для глутаматергических синапсов с VGluT1, существует значительная разница между слоями для глутаматергических синапсов с VGluT2, как с VGluT1, так и с VGluT2, а также для тормозных синапсов. ( C ) Фракция тормозных синапсов в слоях с 1 по 4 соматосенсорной коры мыши. ( D ) Фракция синапсов VGluT2 в слоях с 1 по 4 соматосенсорной коры мыши.
Рисунок 5
Изменения плотности синапсов в…
Рисунок 5
Изменения плотности синапсов по слоям. ( A ) Сводная таблица, показывающая…
Изменения плотности синапсов по слоям. ( A ) Сводная таблица, показывающая типы синапсов, которые имеют значительное увеличение или уменьшение плотности синапсов между мышами дикого типа и мышами с нокаутом. В каждой строке показаны графики процентной разницы в плотности между мышами дикого типа и мышами с нокаутом. Графики имеют цветовую кодировку, чтобы указать направление изменения и статистическую значимость. ( B – D ) Различия в плотности синапсов для разных типов синапсов для трех ожидаемых размеров синапсов.
Рисунок 6
Распределение размеров постсинаптических маркеров. (…
Рисунок 6
Распределение размеров постсинаптических маркеров. ( A ) Сводная таблица, показывающая различия в…
Распределение размеров постсинаптических маркеров. ( A ) Сводная таблица, показывающая различия в распределении размера PSD-95 точек, связанных с синапсами по типу. Основной вывод заключается в том, что для возбуждающих синапсов в целом наблюдается увеличение количества маленьких точек PSD-95 у мышей с нокаутом FMR1, в то время как количество очень больших точек PSD-95, связанных с синапсами, содержащими VGluT2, уменьшается. ( B – E ) Графики, показывающие распределение точек PSD-95 как для мышей дикого типа, так и для мышей с нокаутом. ( F ) Распределение размеров гефириновых точек, связанных с тормозными синапсами. ( G ) На графике показано значительное увеличение соотношения возбуждающих и тормозных синапсов у мышей с нокаутом.
Рисунок 7
Астроцитарная ассоциация синапсов в…
Рисунок 7
Астроцитарная ассоциация синапсов у мышей дикого типа. ( A ) Часть…
Астроцитарная ассоциация синапсов у мышей дикого типа. ( A ) Часть данных о синапсе дикого типа, где синапс выделен белым прямоугольником. Ниже приведены последовательные срезы выделенного синапса, показанные при большем увеличении. ( B ) Доля синапсов, связанных с астроцитами, варьируется в зависимости от типа синапса. ( C ) Показатель взаимной корреляции как функция бокового смещения между двумя каналами. Пары синаптических маркеров VGluT1 — PSD-95 и GAD-гефирин показаны для сравнения. Корреляция между синаптическими маркерами высока без сдвига и резко падает при латеральном смещении, как и ожидалось для тесно коррелированных пресинаптических и постсинаптических маркеров. С другой стороны, GAD и VGluT1 не совместно локализуются, потому что они маркируют разные типы синапсов, и показатель отрицательной колокализации без сдвига постепенно увеличивается до 0 при латеральном смещении. ( D ) Межслойные различия в астроцитарной ассоциации разных типов синапсов.
Рисунок 8
Краткое описание астроцитарного синапса…
Рисунок 8
Резюме различий плотности астроцитарных синапсов у нокаутных мышей. ( А…
Рисунок 8 Резюме различий плотности астроцитарных синапсов у нокаутных мышей. ( A ) Изменения показаны как разница в плотности в процентах между мышами дикого типа и нокаутными мышами. Графики имеют цветовую кодировку, чтобы указать направление изменения и статистическую значимость. Плотность уменьшается между мышами дикого типа и нокаутными мышами для слоев 1–3, и разница особенно заметна для средних и больших синапсов. ( B ) Изменения доли синапсов, прилегающих к астроцитарным отросткам.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Изменения в синапсах гиппокампа CA1 в мышиной модели синдрома ломкой Х-хромосомы.
Джаваид С., Кидд Г.Дж., Ван Дж., Светлик С., Датта Р., Трапп Б.Д. Джаваид С. и др. Глия. 2018 Апрель; 66 (4): 789-800. doi: 10.1002/glia.23284. Epub 2017 23 декабря. Глия. 2018. PMID: 2
- 95
Бесплатная статья ЧВК.
Секретируемый астроцитами тромбоспондин-1 модулирует дефекты синапсов и позвоночника в модели ломких X мышей.
Cheng C, Lau SK, Doering LC. Ченг С и др. Мол Мозг. 2016 авг. 2;9(1):74. doi: 10.1186/s13041-016-0256-9. Мол Мозг. 2016. PMID: 27485117 Бесплатная статья ЧВК.
Селективная делеция астроглиального FMRP нарушает регуляцию транспортера глутамата GLT1 и способствует фенотипам синдрома ломкой Х-хромосомы in vivo.
Хигашимори Х., Шин К.С., Чианг М.С., Морел Л., Шони Т.А., Нельсон Д.Л., Ян Ю. Хигашимори Х. и др. Дж. Нейроски. 2016 6 июля; 36 (27): 7079-94. doi: 10.1523/JNEUROSCI.1069-16.2016. Дж. Нейроски. 2016. PMID: 27383586 Бесплатная статья ЧВК.
Состояние синапсов при синдроме ломкой Х-хромосомы.
Пфайффер Б.Е., Хубер К.М. Пфайффер Б.Э. и соавт. Нейробиолог. 2009 Октябрь; 15 (5): 549-67. дои: 10.1177/1073858409333075. Epub 2009 26 марта. Нейробиолог. 2009. PMID: 1
70 Бесплатная статья ЧВК. Обзор.
Синдром ломкой Х-хромосомы: дисфункция ГАМКергической системы и цепи.
Палушкевич С.М., Мартин Б.С., Хантсман М.М. Палушкевич С.М. и соавт. Дев Нейроски. 2011;33(5):349-64.
дои: 10.1159/000329420. Epub 2011, 21 сентября.
Дев Нейроски. 2011.
PMID: 21934270
Бесплатная статья ЧВК.
Обзор.
Посмотреть все похожие статьи
Цитируется
Новая роль нейрон-глии в неврологических расстройствах: краткий обзор.
Рахман М.М., Ислам М.Р., Ямин М., Ислам М.М., Саркер М.Т., Мим А.Ф.К., Актер А., Эмран Т.Б., Кавалу С., Шарма Р. Рахман М.М. и др. Оксид Мед Селл Лонгев. 2022 22 августа; 2022:3201644. дои: 10.1155/2022/3201644. Электронная коллекция 2022. Оксид Мед Селл Лонгев. 2022. PMID: 36046684 Бесплатная статья ЧВК. Обзор.
Эффективное и точное обнаружение синапсов с помощью микроскопии с селективным структурированным освещением на предполагаемых интересующих областях ультратонких серийных срезов.
Kim GT, Bahn S, Kim N, Choi JH, Kim JS, Rah JC. Ким Г.Т. и др. Фронт Нейроанат. 2021 15 ноя; 15:759816. doi: 10.3389/fnana.2021.759816. Электронная коллекция 2021. Фронт Нейроанат. 2021. PMID: 34867216 Бесплатная статья ЧВК.
Создание модели астроцитов, полученных из плюрипотентных стволовых клеток человека, с идентичностью переднего мозга.
Петери Великобритания, Питконен Дж., Утами К.Х., Паавола Дж., Ройбон Л., Пулади М.А., Кастрен М.Л. Петери Великобритания и др. наук о мозге. 2021 9 февраля; 11 (2): 209. doi: 10.3390/brainsci11020209. наук о мозге. 2021. PMID: 33572154 Бесплатная статья ЧВК.
FMRP регулирует субклеточное распределение плотности корковых дендритных шипов неклеточно-автономным образом.
Бланд К.М., Аарон А., Уайденер Э.Л., Сонг М.И., Кейси З.О., Зуо Ю., Видал Г.С. Бланд К.М. и соавт. Нейробиол Дис. 2021 март; 150:105253. doi: 10.1016/j.nbd.2021.105253. Epub 2021 6 января. Нейробиол Дис. 2021. PMID: 33421563 Бесплатная статья ЧВК.
Взаимодействие нейронов и глии при нарушениях развития нервной системы.
Ким Ю.С., Чхве Дж., Юн Б.Е. Ким Ю.С. и др. Клетки. 2020 сен 27;9(10):2176. doi: 10.3390/cells
- 76.
Клетки. 2020.
PMID: 329
Бесплатная статья ЧВК. Обзор.
использованная литература
- Хантер Дж. и др. Эпидемиология синдрома ломкой Х-хромосомы: систематический обзор и метаанализ. Американский журнал медицинской генетики, часть A. 2014; 164: 1648–1658. doi: 10.1002/ajmg.a.36511.
—
DOI
—
пабмед
- Хантер Дж. и др. Эпидемиология синдрома ломкой Х-хромосомы: систематический обзор и метаанализ. Американский журнал медицинской генетики, часть A.
- Берри-Кравис Э. Эпилепсия при синдроме ломкой Х. Медицина развития и детская неврология. 2002; 44: 724–728. doi: 10.1111/j.1469-8749.2002.tb00277.x. — DOI — пабмед
- Фиш Г. С., Сименсен Р.Дж., Шроер Р.Дж. Продольные изменения показателей когнитивного и адаптивного поведения у детей и подростков с мутацией Fragile X или аутизмом. Журнал аутизма и нарушений развития. 2002; 32: 107–114. дои: 10.1023/A:1014888505185.
—
DOI
—
пабмед
- Фиш Г.
- Пенагарикано О., Мулле Дж.Г., Уоррен С.Т. Патофизиология синдрома ломкой Х-хромосомы. Ежегодный обзор геномики и генетики человека. 2007; 8: 109–129. doi: 10.1146/annurev.genom.8.080706.0. — DOI — пабмед
- Джин П. , Алиш Р.С., Уоррен С.Т. РНК и микроРНК при ломкой Х умственной отсталости. Природа клеточной биологии. 2004; 6: 1048–1053. doi: 10.1038/ncb1104-1048.
—
DOI
—
пабмед
- Джин П.
Типы публикаций
термины MeSH
вещества
Грантовая поддержка
- R01 Mh209475/MH/NIMH NIH HHS/США
- R01 Mh204227/MH/NIMH NIH HHS/США
- R01 NS094499/NS/NINDS NIH HHS/США
- R01Mh204227/США Департамент здравоохранения и здравоохранения Социальные службы | Национальные институты здоровья (NIH)/Международные
- R01 Mh211768/MH/NIMH NIH HHS/США
- R01 NS0/NS/NINDS NIH HHS/США
Многогранное воздействие каменистого коралла Porites astreoides на численность пикопланктона и состав сообщества
- DOI:10. 1002/lno.10389
- Идентификатор корпуса: 55198906
@article{McNally2017MultifacetedIO,
title={Многогранное воздействие каменистого коралла Porites astreoides на численность пикопланктона и состав сообщества},
автор = {Шон П. МакНалли и Рэйчел Дж. Парсонс, Элисон Э. Санторо и Эми Апприлл},
journal={Лимнология и океанография},
год = {2017},
объем = {62}
} - Sean P. McNally, R. Parsons, A. Apprill
- Опубликовано 1 января 2017 г.
- Науки об окружающей среде
- Лимнология и океанография
Пикопланктон способствует существенному повторному использованию питательных веществ в олиготрофных водах, поддерживающих экосистемы коралловых рифов. Несмотря на этот факт, мало данных о том, как специфические взаимодействия между кораллами, планктонными бактериями и археями (пикопланктоном) влияют на динамику питательных веществ и продуктивность рифов. Здесь мы использовали эксперименты с мезокосмом, чтобы исследовать, как кораллы и коралловая слизь влияют на пикопланктон и питательные вещества в рифовых водах. В течение 12 дней мы отслеживали концентрацию питательных веществ…
View on Wiley
aslopubs.onlinelibrary.wiley.comКоралловая экосфера: уникальная среда обитания коралловых рифов, способствующая взаимодействию между кораллами и микробами
- L. Weber, P. Gonzalez-Díaz, M. Armenteros, A. Apprill
Науки об окружающей среде, география
Лимнология и океанография
- 2019
Полевые исследования для сравнения биомассы, разнообразия, состава и функциональной способности микроорганизмов в пробах морской воды небольшого объема, отобранных рядом с пятью видами кораллов с морская вода, собранная на расстоянии более 1 м от рифового субстрата на тех же рифах, обнаружила заметную связь между кораллом Porites astreoides и коралловым симбионтом Endozoicomonas, что предполагает рекрутирование и/или выделение этих клеток в окружающую морскую воду.
Обесцвечивание вызывает гетеротрофию кораллов по диазотрофам и Synechococcus
Показано, что кораллы поглощают больше азота, полученного из планктонных диазотрофов в условиях обесцвечивания, что может иметь серьезные последствия для восстановления обесцвечивания кораллов.
Избирательное поглощение членов пелагического микробного сообщества карибскими рифовыми кораллами
- Kenneth D. Hoadley, Marian I. Hamilton, Camille Poirier, Chang Jae Choi, Cheuk-Man Yung, A. Worden
Науки об окружающей среде
Прикладная и экологическая микробиология
- 2021
Исследование показывает, что бенто-пелагическое взаимодействие посредством селективного выпаса микробных групп влияет на поток углерода и поддерживает неоднородность микробных сообществ, лежащих над коралловыми рифами.
Популяция Диля и функциональная синхронность микробных сообществ на коралловых рифах
- L. Kelly, Craig E. Nelson, F. Rohwer
Науки об окружающей среде
Nature Communications
- 2019
Метагеномный анализ, характеризующий микробные сообщества в толще воды, перекрывающей 16 отдаленных участков переднего рифа в течение дневного цикла, показывает, что состав микробного сообщества больше отличается между дневными и дневными пробами, собранными в одном и том же месте, чем между дневные или ночные пробы, собранные на географически удаленных рифах.
Динамика симбионтов бактерий и водорослей у ранних рекрутов, подвергшихся воздействию двух взрослых видов кораллов
- K. Damjanovic, L. Blackall, Patricia Menéndez, M. V. van Oppen
Environmental Science
Coral Reefs
- 2019
It is indicated that Pocillopora acuta recruits harbor dynamic and diverse bacterial assemblages, which were not под влиянием близлежащих взрослых кораллов в экспериментальном дизайне авторов.
Стратегии усвоения азота диазотрофами различаются в зависимости от вида кораллов-склерактиний
- V. Meunier, S. Bonnet, F. Houlbrèque
Науки об окружающей среде
Frontiers in Marine Science
- 2021
Рифообразующие кораллы обычно процветают в бедных питательными веществами тропических водах, где помимо других элементов наличие азота (N) часто ограничивает первичную продуктивность. В дополнение к их тесной связи…
Микробные сообщества коралловых рифов Красного моря
- M. Neave, A. Apprill, G. Aeby, S. Miyake, C. Voolstra
Науки об окружающей среде
Коралловые рифы Красного моря
- 2019
Несмотря на сильные экологические экстремальные явления, стимулирующие существование свободноживущих микробных сообществ в Красном море, микробы в жестко регулируемой симбиотической среде, по-видимому, сохраняются, хотя специализация на уровне штаммов и генотипов сохраняется. области продолжающихся исследований.
Численность, разнообразие и функциональные возможности планктонных бактерий и микроэукариот в системе коралловых рифов островов Сиша, Китай
- Xiaoli Zhang, Lei Wang, Jianmin Zhao
Науки об окружающей среде
Journal of Ocean University of China
- 2022
-системы рифов не получили должного внимания. Мы…
Связь реакции фотоакклиматизации и изменений микробиома между родственными видами рифовых кораллов, сегрегированными по глубине
Это исследование предполагает, что эти родственные виды адаптированы к различным световым условиям, частично обусловленным способностью водорослей к фотоакклиматизации и устойчивостью микробиома, подчеркивая важность специализации ниш у симбиотических кораллов для поддержания видового разнообразия.
Микробные сигнатуры охраняемых и затронутых рифов Северной Карибского моря: переход от Кубы к Флорида-Кис
- L. Weber, P. González-Díaz, A. Apprill
Науки об окружающей среде
Микробиология окружающей среды
- 2019
Предполагается, что защита и прибрежная природа Хардинес-де-ла-Рейна могут сохранить олиготрофную парадигму и метаболическую зависимость сообщества от первичной продукции пикоцианобактерий.
ПОКАЗАНЫ 1-10 ИЗ 115 ССЫЛОК
СОРТИРОВАТЬ ПОРелевантности Наиболее влиятельные документыНедавность
Экологические выводы из глубокого скрининга сложных бактериальных консорциумов, связанных с кораллом, Porites astreoides
- M. Rodriguez-Lanetty, Camila Granados-Cifuentes, A. Barberán, Anthony J. Bellantuono, C. Bastidas
Environmental Science, Geography
Molecular ecology
- 2013
This study focused on the exploration ассоциаций микробных сообществ, связанных с экологически важным карибским склерактиновым кораллом Porites astreoides, с помощью высокопроизводительного секвенирования Illumina фрагмента V5 гена 16S рРНК для выявления специфического паттерна бактериальной ассоциации, появляющейся для этого вида кораллов и для многих других кораллов в пределах род Пориты.
Метагеномный анализ микробного сообщества, ассоциированного с кораллом Porites astreoides.
- L. Wegley, R. Edwards, Beltran Rodriguez-Brito, Hong Liu, F. Rohwer
Науки об окружающей среде, биология
Микробиология окружающей среды
- 2007
Экссудаты кораллов и макроводорослей различаются по составу нейтральных сахаров и по-разному обогащают линии рифового бактериопланктона
Показано, что экссудаты водорослей обогащены компонентами DCNS, способствуют быстрому росту бактериопланктона и отбирают бактериальные популяции с более потенциальными VF, чем коралловые экссудаты.
Локальная геномная адаптация микробиомов коралловых рифов к градиентам естественной изменчивости и антропогенным стрессорам
- L. Kelly, Gareth J. Williams, F. Rohwer
Науки об окружающей среде, биология
Proceedings of the National Academy of Sciences
- 2014
Предполагается, что первичные источники углерода определяют структуру сообщества и что локальная биогеохимия определяет более мелкие метаболические функции, а основные таксоны адаптируются к местным условиям путем отбора выгодных метаболических генов.
Разнообразие и распространение бактерий, ассоциированных с кораллами
- F. Rohwer, V. Seguritan, F. Azam, N. Knowlton
Науки об окружающей среде
- 2002
Анализ ветвящихся кораллов Porites furcata показал, что бактериальные риботипы также могут быть пространственно структурированы внутри колоний, а кораллы и рифы представляют собой ландшафты разнообразных, экологически структурированных прокариотических сообществ.
Удаление частиц сообществами коралловых рифов: пикопланктон является основным источником азота
- M. Ribes, R. Coma, M. Atkinson, R. Kinzie
Науки об окружающей среде
- 2003
В целом удаление азота из частиц похоже по величине аналогично удалению растворенного неорганического азота: таким образом, пикопланктон является основным источником азота для этих сообществ коралловых рифов.
Круговорот азота в кораллах: ключ к пониманию функционирования холобионтов?
- N. Rädecker, C. Pogoreutz, C. Voolstra, J. Wiedenmann, C. Wild
Науки об окружающей среде
Тенденции в микробиологии
- 2015
Доказательства повышенной микробной активности в интерстициальном пространстве разветвленных кораллов: возможные последствия для метаболизма кораллов -, Nh5+, реактивного молибдатного фосфора, бактериальной и жгутиковой биомассы и растворенного органического вещества и по сравнению с концентрациями в окружающей воде.
Удаление бактерий и динамика питательных веществ в пределах кораллового рифа Кюрасао (Нидерландские Антильские острова)
- S. Scheffers, G. Nieuwland, R. Bak, F.C.V. Duyl
Науки об окружающей среде
Coral Reefs
- 2004
(DSi) и источник NOx, что предполагает сильную реминерализацию и нитрификацию в полостях с потенциальной ролью губчато-симбиотической микробной нитрификации.
Реакция микробного сообщества на нерест кораллов в лагунах и рифовых равнинах на Гавайях, США
Различия в составе сообщества бактериопланктона, клеточном обилии микроорганизмов и скорости бактериальной продукции были очевидны между лагуной и рифовыми участками в период отсутствия нереста, что, вероятно, свидетельствует о важности режима поверхностного стока для микробных сообществ прибрежных рифов. .
Многогранный подход Джудит Шехтер к самовыражению
Источник изображения: Фрагмент фильма «Дикий ребенок» (2012), Джудит Шехтер.
Доказательства того, что мультимедийная художница Джудит Шехтер десятилетиями работала с инструментами компьютерного дизайна, может быть трудно определить.
Ее витражные композиции богато украшены, полны воображения и искусно детализированы вручную. Там, где кончается физическое и начинается цифровое, только добавляет сказочности, неземности ее работам — обновлению старых условностей витражей. Сочетание Шехтера этой традиционной формы искусства с современными темами и предметами реконтекстуализирует и то, и другое, что является частью большой и растущей тенденции дизайна, которую Adobe Stock назвал «строгим романтизмом».
Для некоторых художников самовыражение может проявляться как индивидуальный акт автопортрета, но для Шехтера есть желание выразить глубоко личное через универсальное.
Источник изображения: «Ты здесь» (2008), Джудит Шехтер.
«Вам нужно понять, что в вас такого уникального, а также что делает вашу работу невероятно универсальной», — говорит Шехтер.
Что выделяет работы Шехтер, так это ее непрерывные эксперименты в среде, уходящей своими корнями в многовековое искусство Средневековья и Ренессанса. Она также опирается на свои пожизненные отношения с конкретными картинами, созданными на протяжении всей истории искусства, влияние которых очевидно даже через слои абстракции, которые Шехтер вплетает в свои убедительные современные работы из стекла.
Персонажи, которые Шехтер изображает в своих изделиях из стекла, представляют собой серию красивых, захватывающих и сложных фигур в столь же сложных ситуациях. Одинокие субъекты — это почти всегда женщины, вовлеченные в акты самосознания, тайные размышления или неясные ритуалы. Ее более крупные работы часто представляют собой босхианские сценарии веселья и хаоса с широким спектром персонажей, действий и взглядов.
Источник изображения: «В плену наводнения» (1990), Джудит Шехтер.
От физического к Photoshop
Кажется нелогичным думать, что процесс Шехтера, который начинается с нарисованных от руки эскизов и заканчивается часами тщательной гравировки и шлифовки цветного стекла, имеет какое-либо отношение к современным технологиям — и, много лет этого не было. Но в 2000 году, уже спустя пару десятилетий своей художественной карьеры, Шехтер произвела революцию в своем композиционном процессе, поскольку ее отношение к технологиям начало меняться.
«Раньше недалеко от того места, где я жил в то время, был центр Xerox, который назывался Can-Do, и я приносил свои чертежи в Can-Do и ксерокопировал их во всех размерах», — говорит Шехтер. «Мой счет за Xerox составлял сотни долларов в месяц или что-то в этом роде». Шехтер ранее пробовала свои силы в технических инструментах с помощью подержанного Apple 2, который требовал ручного кодирования для работы с основными программами. Все пошло не так, как планировалось.
«Я выбросил его в окно, — говорит Шехтер. «Я не получил заметки о том, что операционные системы становятся удобными для пользователя — я просто не обращал на это внимания, — но я решил, что мне нужно купить машину Xerox, потому что это было сумасшествием. Итак, мой маленький мозг как горошина перемалывался, и я подумал, может быть, если у меня будет компьютер, я смогу организовать на нем свой список рассылки. И, по сути, компьютер — это была такая же цена, как покупка ксерокса, поэтому я подумал, хорошо, я куплю компьютер, и у меня есть сканер».
Источник изображения: «Натюрморт с грабителем банка» (1996), Джудит Шехтер.
Примерно в то же время Шехтер прочитал статью иллюстратора Марка Райдена, в которой он обсуждал преимущества сканирования в своих эскизах. Затем Райден смог «опробовать бесчисленное количество миллионов макетов за один день, по сути, просто перемещая элементы в документе Photoshop», — вспоминает Шехтер. «И я подумал: «Ах! Да! Это то, что я хочу делать!» И это то, что я сейчас делаю».
Она начала с сокращенной версии Photoshop, прилагаемой к ее сканеру, и программа тут же дала откровение.
«Я чувствовал, что Photoshop был моим старым другом, — говорит Шехтер. «Я человек, который нажимает на каждую кнопку, поэтому сначала я все разрушил, а потом мне пришлось все чинить». В результате Шехтер быстро включила Photoshop в свой художественный рабочий процесс.
«Вот что я вам скажу, я очень люблю Adobe Photoshop — он изменил весь мой взгляд на искусство в очень, очень положительную сторону», — говорит она. «Мне всегда жаль людей, которых это пугает. Это невероятно удобно, если вы художник. Я чувствую, что это, должно быть, было сделано художниками для художников».
Источник изображения: «Дитя-невеста» (2001), Джудит Шехтер.
Искусство в жизни — и жизнь в искусстве
Баланс между специфичностью и универсальностью в творчестве Шехтер затрагивает самую суть уязвимости, связанной с видимостью — или невидимостью — женской формы, особенно несовершенной или стареющей.
«Я боролся со своим телом, как и многие, многие, многие люди до меня, — говорит Шехтер, — и я думаю, что это проявляется в моей работе». Фигуры Шехтер часто имеют видимые вены под кожей или преувеличенные черты — акцент на естественных аспектах тела, которые женщины часто вынуждены скрывать, — и она описывает их как попытку выразить несоответствие между «то, что мы видим, и тем, о чем мы думаем». человеческие тела».
Субъекты Шехтера часто кажутся захваченными борьбой между собственной святостью себя и требованиями более могущественных внешних сил, воздействующих на них.
«Они не я, — говорит Шехтер о своих фигурах, — они как прокси, кукольная версия меня, и я хочу, чтобы они были лучше меня и красивее меня, чтобы компенсировать мое тело». и лицевая дисморфия.
«Но я не могу сделать их слишком красивыми, — смеется она, — потому что тогда я больше не могу с ними себя идентифицировать!»
Последние новости о многогранной пользе для здоровья, связанной с имбирем и его биологически активными компонентами
У вас не включен JavaScript. Пожалуйста, включите JavaScript чтобы получить доступ ко всем функциям сайта или получить доступ к нашему страница без JavaScript.
Выпуск 2, 2021 г.
Из журнала:
Еда и функции
Недавнее обновление о многогранной пользе для здоровья, связанной с имбирем и его биологически активными компонентами
.Рун-Хуэй Ма, аб Чжи-Цзин Ни, аб Юн-Ян Чжу, аб Киран Такур, аб Поклонник Чжан, и Юань-Юань Чжан, и Фэй Ху, аб Цзянь-Го Чжан* аб а также Чжао-Цзюнь Вэй * абв
Принадлежности автора
* Соответствующие авторы
и Школа биологических наук и инженерии, Совместный инновационный центр производства и безопасности продуктов питания, Северный университет Миньцзу, Иньчуань 750021, Китайская Народная Республика
Электронная почта: 1120355093@qq. com, [email protected], [email protected], [email protected], [email protected], [email protected], [email protected], [email protected], zjwei@hfut. edu.cn
б Школа пищевой и биологической инженерии, Технологический университет Хэфэй, Хэфэй 230009, Китайская Народная Республика
с Колледж биологических наук и технологий Университета Фучжоу, Фучжоу, Китайская Народная Республика
Аннотация
3043 in vitro , in vivo и клеточные линии). В этом обзоре мы также сосредоточились на терапевтических эффектах имбиря и его компонентов благодаря их эффективным антиоксидантным свойствам. Их потребление может уменьшить или задержать прогрессирование сопутствующих заболеваний, таких как рак, диабет и ожирение, посредством модуляции генетической и метаболической активности. Обновленные данные могут прояснить связь процессов экстракции с составляющими и биологическими проявлениями. Мы собрали современные знания (включая последние клинические данные) о биологически активных соединениях и биоактивности имбиря. Также обсуждаются их подробные механизмы, которые могут заложить основу для их пищевых и медицинских применений.- Эта статья является частью тематического сборника: Последние обзорные статьи
Варианты загрузки Пожалуйста, подождите. ..
Информация о товаре
- ДОИ
- https://doi.org/10.1039/D0FO02834G
- Тип изделия
- Обзор статьи
- Отправлено
- 29 окт 2020
- Принято
- 03 дек. 2020
- Впервые опубликовано
- 08 дек. 2020
Скачать цитату
Функц. , 2021 , 12 , 519-542
BibTexEndNoteMEDLINEProCiteReferenceManagerRefWorksRIS
Разрешения
Запросить разрешения
Социальная деятельность
Получение данных из CrossRef.